Regulatory Mechanisms (OCR A-Level Biology A): Revision Notes
Regulatory Mechanisms
Gene expression in cells is controlled through multiple regulatory mechanisms operating at different stages. These mechanisms determine when and where specific genes are activated, ensuring that proteins are produced only when needed. Three main levels of control exist: transcriptional, post-transcriptional, and post-translational.
Each eukaryotic cell contains a complete genome with all chromosomal information. However, not all genes are active simultaneously. Structural genes code for functional proteins such as enzymes, membrane carriers, and hormones. Separate regulatory genes produce proteins and RNA molecules that control structural gene expression at the three levels mentioned above.
Control at the transcriptional level
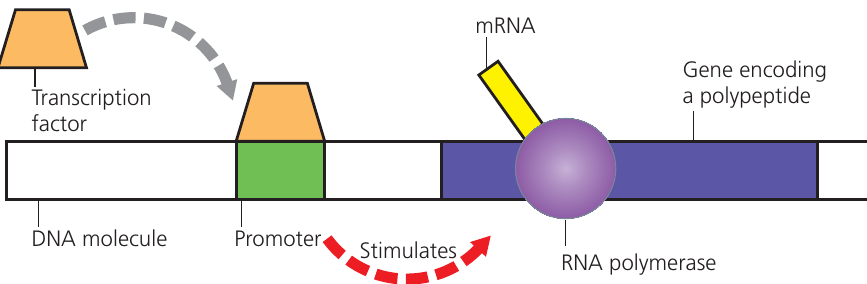
Transcription factors and promoter regions
Transcription — the process of copying DNA sequences into mRNA — does not occur for all genes simultaneously. Cells transcribe only those genes coding for currently required proteins, except in cases of regulatory errors such as in cancer cells.
Each gene has an associated promoter region, a DNA sequence typically located approximately base pairs upstream from the gene start site. Transcription begins when transcription factors (regulatory proteins) bind to the promoter region. This binding allows RNA polymerase to attach and initiate mRNA synthesis.
Oestrogen regulation of gene expression
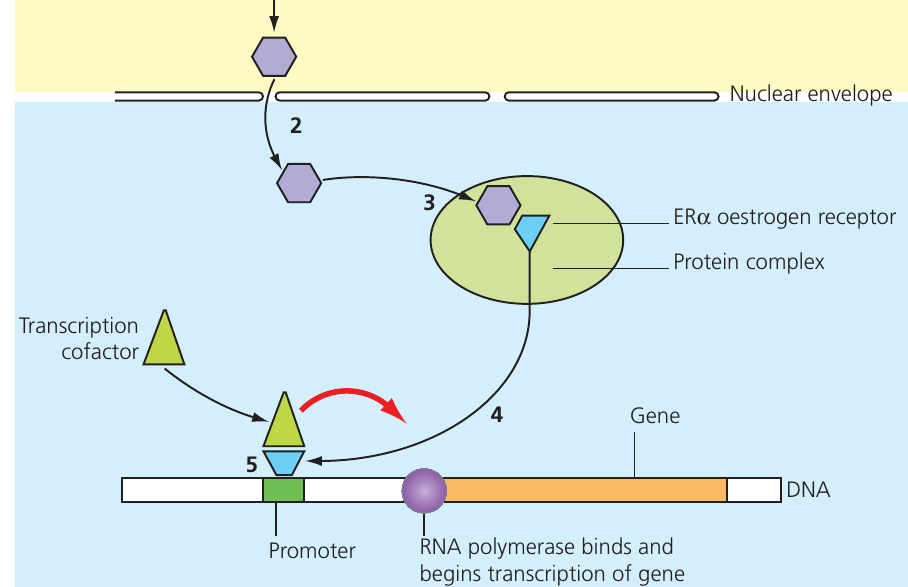
The hormone oestrogen regulates the oestrus cycle and sperm production in mammals. Being lipid-soluble, oestrogen diffuses through plasma membranes and enters the nucleus, where it binds to oestrogen receptors (ERα). These receptors function as transcription factors capable of initiating transcription for up to different genes.
Worked Example: The Oestrogen Regulatory Pathway
The regulatory process follows these steps:
- Oestrogen diffuses through the plasma membrane and nuclear envelope
- It binds to an ERα oestrogen receptor within a protein complex in the nucleus
- Binding causes a conformational change in the receptor
- The receptor dissociates from the inhibitory protein complex
- The activated receptor binds to the promoter region of target genes
- Additional transcription cofactors are recruited
- RNA polymerase can now bind and transcribe the gene
This demonstrates how lipid-soluble hormones can directly influence gene expression by acting as transcription factors.
The lac operon in prokaryotes
Some genes are constitutive — transcribed continuously because their products are always needed. Others encode inducible enzymes that are produced only when required, preventing wasteful protein synthesis.
The lac operon in Escherichia coli exemplifies transcriptional control. This DNA section contains three structural genes and a shared promoter, regulating enzymes for lactose metabolism. The system ensures these proteins are synthesized only when lactose is present and glucose levels are low.
Key Term: Operon
An operon is a functional DNA unit consisting of multiple structural genes controlled by one promoter. This organization allows bacteria to coordinate the expression of related genes efficiently.
Components of the lac operon
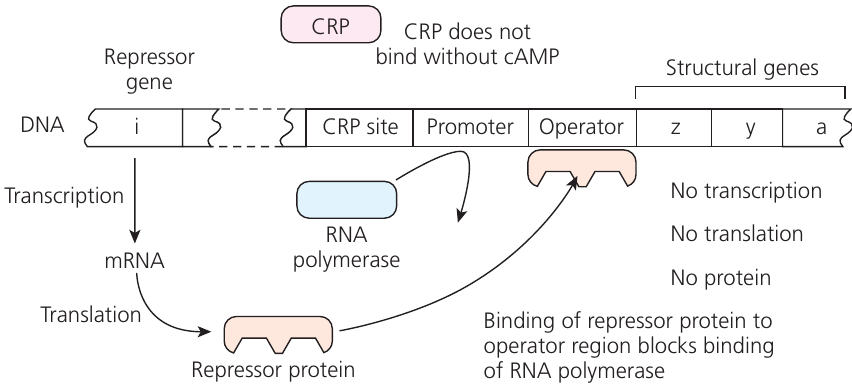
The lac operon consists of several key components, each playing a specific role in the regulatory mechanism:
CRP site
The CRP (cyclic AMP receptor protein) site is the binding location where the CRP-cAMP complex attaches to facilitate RNA polymerase binding to the promoter. CRP can only bind when complexed with cyclic AMP (cAMP). High glucose levels reduce cAMP concentration, preventing CRP binding and keeping the operon inactive.
Promoter region
The promoter region serves as the RNA polymerase attachment site for transcription initiation. When blocked by repressor protein, RNA polymerase cannot bind, preventing mRNA production and subsequent protein synthesis.
Operator region
The operator region is located adjacent to structural genes and functions as the repressor protein binding site. It acts as a molecular switch controlling gene transcription — when repressor binds, transcription and translation are blocked.
Structural genes
Three genes code for β-galactosidase, lactose permease, and another metabolic enzyme. All are transcribed together when the operon is active. Each mRNA section has its own ribosomal binding site for independent translation.
Repressor gene
The repressor gene is located elsewhere on bacterial DNA, not part of the operon itself. It codes for a repressor protein that binds tightly to the operator, causing DNA loop formation that prevents RNA polymerase binding. This maintains the operon in an inactive state by default.
Operon function: absence of lactose
Example: Lac Operon with High Glucose, No Lactose
When glucose is abundant and lactose absent, the operon remains inactive through the following mechanism:
- High glucose levels keep cAMP concentration low
- CRP cannot bind to the CRP site without cAMP
- The repressor protein binds to the operator region
- RNA polymerase cannot access the promoter
- No transcription occurs
- No mRNA is produced
- The three enzymes are not synthesized
Result: Energy conservation — the cell doesn't waste resources producing unnecessary enzymes.
Operon function: presence of lactose
Example: Lac Operon with Low Glucose, Lactose Present
When glucose is scarce and lactose present, the operon activates through this process:
- Low glucose levels increase cAMP concentration
- CRP-cAMP complex binds to the CRP site
- Lactose acts as an inducer, binding to the repressor protein
- Binding changes the repressor's shape, preventing operator attachment
- The repressor releases from the operator region
- RNA polymerase can now bind to the promoter
- All three structural genes are transcribed into a single mRNA molecule
- Each gene section is independently translated
- The three enzymes for lactose metabolism are produced
Result: Efficient lactose metabolism — the cell produces the necessary enzymes only when they're needed.
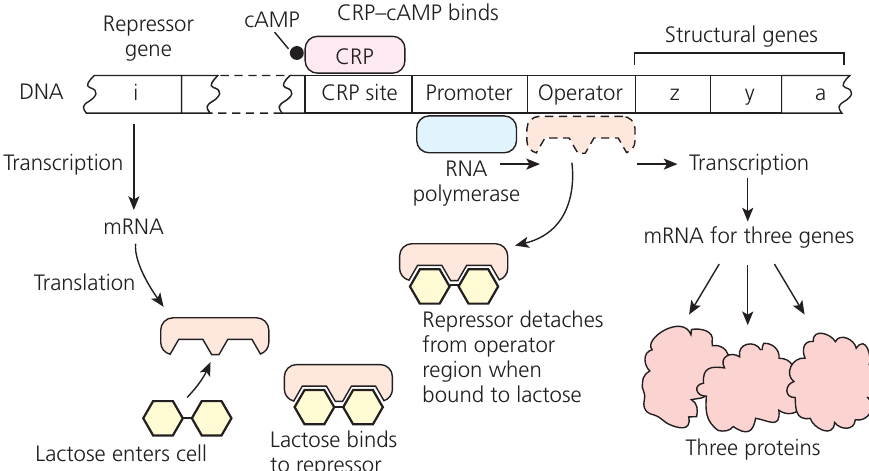
This dual control mechanism — requiring both lactose presence and glucose absence — ensures efficient energy utilization by the bacterial cell. Glucose is the preferred energy source, so the operon only activates when glucose is unavailable.
Control at the post-transcriptional level
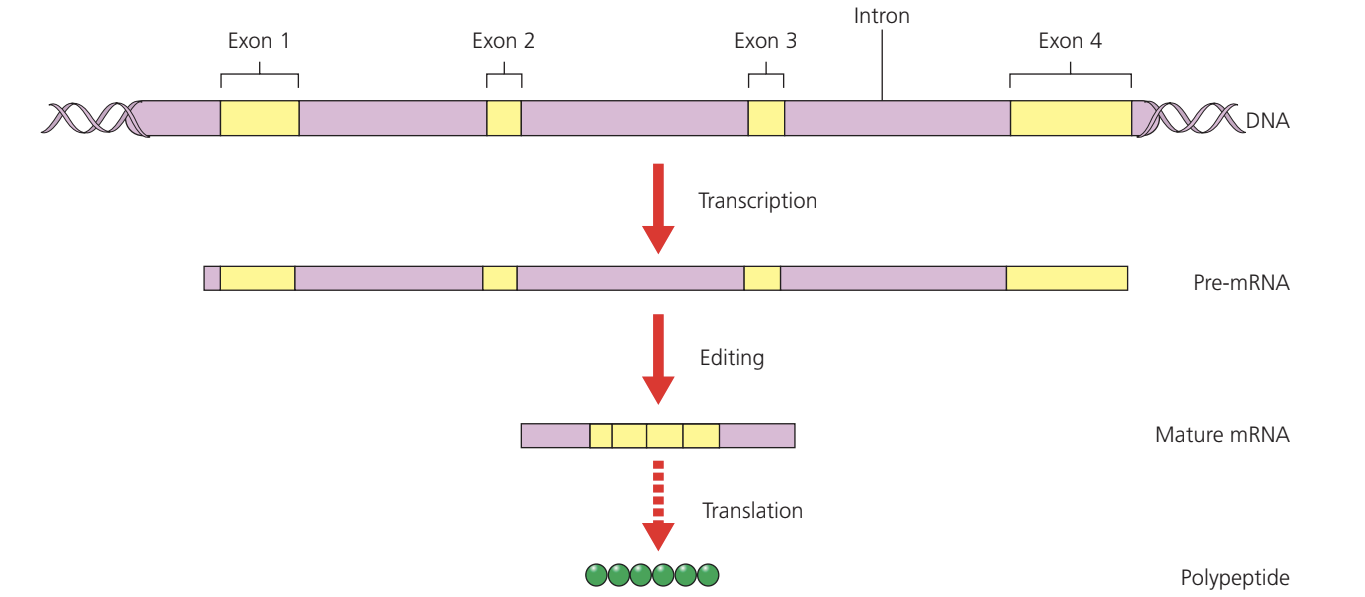
Eukaryotic genomes contain substantial non-coding DNA, including tandem repeat sequences between genes that are not transcribed. Additionally, genes themselves contain non-coding regions that are transcribed but later removed.
Introns and exons
Genes consist of alternating coding and non-coding sections:
- Exons are coding sequences that encode polypeptides
- Introns are non-coding sequences situated between exons
- During transcription, both are copied into primary mRNA (pre-mRNA)
Before leaving the nucleus, primary mRNA undergoes editing to remove introns. This processing is essential — if introns remained in the mature mRNA, the resulting protein would be non-functional.
mRNA splicing
Small nuclear ribonucleoproteins (snRNPs), pronounced "snurps", are RNA-protein complexes that catalyze mRNA processing. The RNA component cuts and splices the mRNA molecule, removing introns and joining exons to create mature mRNA.
Why Intron Removal is Critical
This editing ensures:
- Only coding sequences are translated into protein
- No amino acids are wasted on non-functional sequences
- Proper gene expression occurs
Without this process, the translated proteins would be completely non-functional, as the intron sequences would disrupt the correct amino acid sequence.
Interestingly, some removed introns are further processed to generate non-coding RNA molecules that may regulate other genes.
Memory Aid for Introns vs. Exons
- Introns are intervening sequences (between coding regions)
- Exons are expressed sequences (translated into protein)
This simple mnemonic helps you remember which sequences are kept and which are removed during mRNA processing.
Control at the post-translational level
Following translation, polypeptides undergo modification in the Golgi apparatus or cytosol. These modifications activate or regulate the final protein products.
Cyclic AMP (cAMP) regulation
Cyclic AMP is an ATP derivative produced by the enzyme adenyl cyclase. It functions as a second messenger in cells, regulating numerous processes:
- Hormone activation
- Cellular transport mechanisms
- Ion channel activation and regulation
- Protein kinase activation
In eukaryotic cells, cAMP activates protein kinase A (PKA), an initially inactive precursor enzyme. Once activated, PKA can activate additional proteins, creating a regulatory cascade.
Allosteric regulation example
Example: Glycogen Phosphorylase Allosteric Regulation
The enzyme glycogen phosphorylase releases glucose from glycogen when muscle cells require energy. This enzyme demonstrates allosteric regulation:
Step 1: cAMP binds to the enzyme's allosteric site (a regulatory site distinct from the active site)
Step 2: Binding induces a conformational change, exposing the active site
Step 3: The enzyme becomes active and breaks down glycogen to glucose
Step 4: ATP and glucose-6-phosphate inhibit the enzyme
Result: Energy-responsive regulation — glycogen breakdown occurs only when cellular energy levels are low, preventing wasteful catabolism when energy is abundant.
Key Term: Allosteric Site
An allosteric site is a regulatory binding location on an enzyme separate from the active site, used by activators or non-competitive inhibitors to modulate enzyme activity. This allows for sophisticated control of enzyme function without blocking the active site directly.
Remember!
Key Points to Remember:
-
Three levels of control regulate gene expression: transcriptional (controlling mRNA synthesis), post-transcriptional (editing mRNA), and post-translational (modifying proteins).
-
Transcription factors bind to promoter regions to initiate gene transcription; the oestrogen receptor exemplifies hormone-mediated transcriptional control.
-
The lac operon demonstrates bacterial gene regulation through a dual control system requiring lactose presence and glucose scarcity for activation.
-
Introns (non-coding sequences) are removed from pre-mRNA by snRNPs during splicing, leaving only exons (coding sequences) in mature mRNA.
-
Cyclic AMP acts as a crucial second messenger in post-translational control, activating enzymes like protein kinase A through allosteric regulation.