Haemoglobin and the Transport of Oxygen and Carbon Dioxide (OCR A-Level Biology A): Revision Notes
Haemoglobin and the Transport of Oxygen and Carbon Dioxide
Introduction
Oxygen has relatively low solubility in water. If oxygen were transported only as dissolved gas in blood plasma, the quantity delivered to respiring tissues would be insufficient to meet metabolic demands. Red blood cells contain haemoglobin, a specialized respiratory pigment that binds oxygen reversibly and transports it efficiently throughout the body. Haemoglobin also contributes to carbon dioxide transport.
The low solubility of oxygen in water is the key reason why complex organisms need specialized oxygen transport molecules. Without haemoglobin, our blood could only carry about 2% of the oxygen we actually need!
Structure of haemoglobin
Haemoglobin is a globular protein with quaternary structure, meaning it consists of multiple polypeptide subunits folded together.
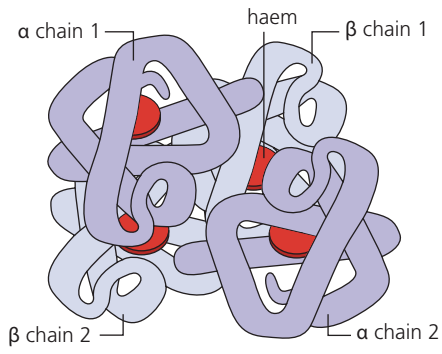
The molecule comprises four polypeptide chains:
- Two α (alpha) chains
- Two β (beta) chains
Each polypeptide chain is associated with a haem group—a prosthetic group containing an iron (Fe²⁺) ion at its centre. The iron ion in each haem group can bind one oxygen molecule. Therefore, a single haemoglobin molecule can bind four oxygen molecules in total.
The "Four and Four" Rule:
- 4 polypeptide chains
- 4 haem groups (one per chain)
- 4 oxygen molecules can be carried
Each Fe²⁺ ion in the haem groups is the actual binding site for O₂.
Haemoglobin exists in two forms:
- Oxyhaemoglobin: haemoglobin bound to oxygen (Hb(O₂)₄)
- Deoxyhaemoglobin: haemoglobin without oxygen (Hb)
Transport of oxygen
Affinity and partial pressure
The affinity of haemoglobin for oxygen refers to its tendency or attraction to bind oxygen molecules. This affinity is not constant—it varies with the partial pressure of oxygen (pO₂) in the surrounding environment.
Partial pressure is the pressure exerted by one gas in a mixture of gases, measured in kilopascals (kPa) or millimetres of mercury (mmHg). As the partial pressure of oxygen increases, haemoglobin's affinity for oxygen increases.
Cooperative binding
When the first oxygen molecule binds to one haem group, it induces a conformational change in the haemoglobin molecule. This structural alteration makes it easier for the second oxygen molecule to bind. The process continues with the third and fourth oxygen molecules, progressively increasing the ease of binding. This phenomenon is called cooperative binding or positive cooperativity.
Think of cooperative binding like opening a door: the first person has to push hard to get it open, but once it's moving, it becomes easier for the next people to go through. Each oxygen molecule that binds makes it easier for the next one to attach!
Oxygen dissociation curve
The relationship between oxygen partial pressure and the percentage saturation of haemoglobin is represented graphically by the oxygen dissociation curve.
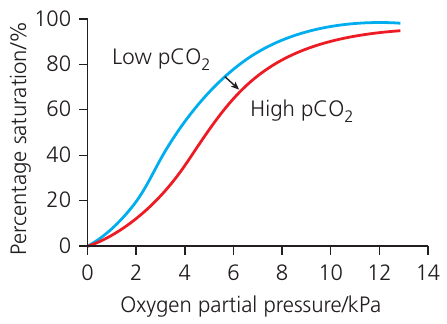
The curve has a characteristic S-shape (sigmoidal), which reflects the cooperative binding mechanism:
- At low pO₂ (0–2 kPa): the curve rises slowly because binding the first oxygen molecule is relatively difficult
- At intermediate pO₂ (2–8 kPa): the curve rises steeply due to cooperative binding making subsequent oxygen binding easier
- At high pO₂ (above 10 kPa): the curve plateaus as haemoglobin approaches 100% saturation
Functional significance
The oxygen dissociation curve demonstrates why haemoglobin is ideally suited for oxygen transport:
In the lungs:
- Partial pressure of oxygen is high (~13 kPa)
- Haemoglobin has high affinity for oxygen
- Haemoglobin becomes ~97% saturated with oxygen
- Oxygen loading is maximized
In respiring tissues:
- Partial pressure of oxygen is low (~4 kPa), varying with activity level
- Haemoglobin has lower affinity for oxygen
- The steep part of the curve means small decreases in pO₂ cause large oxygen release
- Oxyhaemoglobin readily dissociates, delivering oxygen where needed
Why the S-shape is crucial:
The sigmoidal curve ensures:
- Efficient loading - The plateau region means even if lung pO₂ varies slightly, haemoglobin remains nearly fully saturated
- Efficient unloading - The steep middle section means small drops in tissue pO₂ cause large amounts of oxygen to be released
- Reserve capacity - Even after delivering oxygen, haemoglobin retains some O₂ as a safety margin
Bohr effect
The functionality of haemoglobin is enhanced by the Bohr effect, discovered by Christian Bohr. This effect describes how carbon dioxide concentration affects oxygen binding.
When carbon dioxide levels increase (as occurs in actively respiring tissues), the oxygen dissociation curve shifts to the right. This rightward shift indicates that haemoglobin's affinity for oxygen decreases at any given partial pressure of oxygen, promoting oxygen release.
Mechanism: The Bohr effect results from both:
- The direct effect of carbon dioxide on haemoglobin structure
- The decrease in pH caused by carbonic acid formation (CO₂ + H₂O → H₂CO₃)
Physiological advantage: Actively respiring tissues:
- Consume oxygen rapidly (low pO₂)
- Produce carbon dioxide (high pCO₂)
- Have lower pH due to carbonic acid
These conditions work together to maximize oxygen unloading precisely where oxygen demand is greatest. The Bohr effect ensures that haemoglobin releases even more oxygen in metabolically active regions than would occur based on low pO₂ alone.
Remember: "CO₂ Boots Oxygen Off"
Increased CO₂ literally kicks oxygen off haemoglobin, ensuring oxygen is released where cells are actively respiring and need it most. This is a perfect example of how the body's transport systems are intelligently designed for efficiency!
Fetal haemoglobin
In mammals, the developing fetus obtains oxygen from maternal blood via the placenta. However, the partial pressure of oxygen in the placenta is much lower than in adult lungs, potentially creating difficulty for oxygen uptake.
This challenge is overcome through a structural difference in fetal haemoglobin. Instead of two beta (β) chains, fetal haemoglobin contains two gamma (γ) chains. This substitution increases the molecule's affinity for oxygen.
The dissociation curve for fetal haemoglobin lies to the left of the adult curve, meaning fetal haemoglobin achieves higher saturation at lower partial pressures of oxygen. Consequently, at the pO₂ found in the placenta, fetal haemoglobin has greater affinity than maternal haemoglobin, enabling efficient oxygen transfer from mother to fetus.
Why fetal haemoglobin is essential:
At the placental pO₂, both maternal and fetal haemoglobin are present in the same environment. The higher affinity of fetal haemoglobin means oxygen will preferentially bind to the fetal version, allowing oxygen to transfer from mother to baby against a concentration gradient.
After birth, over the first few months of life, fetal haemoglobin is gradually replaced by the adult form as the baby begins breathing air directly.
Physiological adaptations to high altitude
At high altitude, atmospheric pressure is lower, resulting in reduced partial pressure of oxygen. The body responds to chronic low oxygen availability through several adaptations.
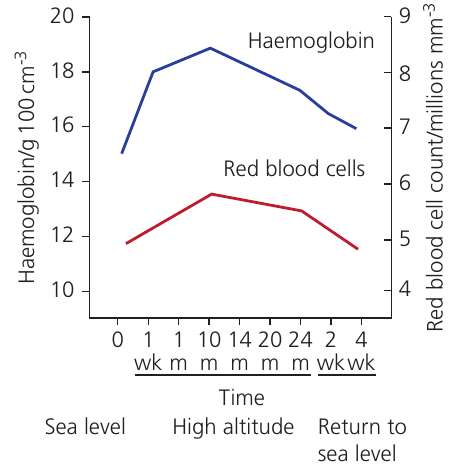
Study: Soldiers at High Altitude (Two-Year Study)
Researchers tracked physiological changes in soldiers stationed at high altitude over two years:
During altitude exposure:
- Haemoglobin concentration increased from ~12 to ~19 g/100 cm³
- Red blood cell count increased from ~5 to ~6.5 million/mm³
- Both parameters peaked around 10–14 months
- Pulse rate, breathing rate, and blood pressure all increased
After returning to sea level:
- Haemoglobin concentration and RBC count gradually decreased
- Values returned toward baseline over several weeks
Analysis: The increased haemoglobin concentration and red blood cell numbers enhance the oxygen-carrying capacity of blood. This compensates for the lower oxygen availability in the air, ensuring adequate oxygen delivery to tissues despite reduced pO₂.
The leveling off after 10 months likely represents a new physiological steady state—a balance between the stimulus for increased production and regulatory mechanisms preventing excessive increases.
Transport of carbon dioxide
Carbon dioxide is more soluble in water than oxygen, but it is still transported in blood through three distinct mechanisms.
1. Dissolved carbon dioxide (approximately 5%)
A small proportion of carbon dioxide dissolves directly in blood plasma and is carried in solution. Carbon dioxide's greater solubility compared to oxygen makes this method more effective for CO₂ than it would be for O₂, though it still accounts for only about 5% of total transport.
2. Hydrogen carbonate ions (70–90%)
This is the principal form of carbon dioxide transport. Hydrogen carbonate (bicarbonate) ions (HCO₃⁻) are formed inside red blood cells but transported in the plasma. This method accounts for 70–90% of all CO₂ transport.
3. Carbaminohaemoglobin
Carbon dioxide can bind directly to haemoglobin, forming carbaminohaemoglobin. Unlike oxygen, which binds to the iron in haem groups, carbon dioxide combines with terminal amino groups on the polypeptide chains of the haemoglobin molecule.
Mnemonic for CO₂ transport forms: "5-7-Rest"
- 5% dissolved in plasma
- 70-90% as bicarbonate (HCO₃⁻)
- Rest as carbaminohaemoglobin
The majority of CO₂ travels as hydrogen carbonate ions because this form is highly soluble and doesn't affect blood pH as much as dissolved CO₂ would.
Mechanism of carbon dioxide transport in red blood cells
The conversion of carbon dioxide to hydrogen carbonate ions involves several coordinated steps that occur simultaneously throughout the body.
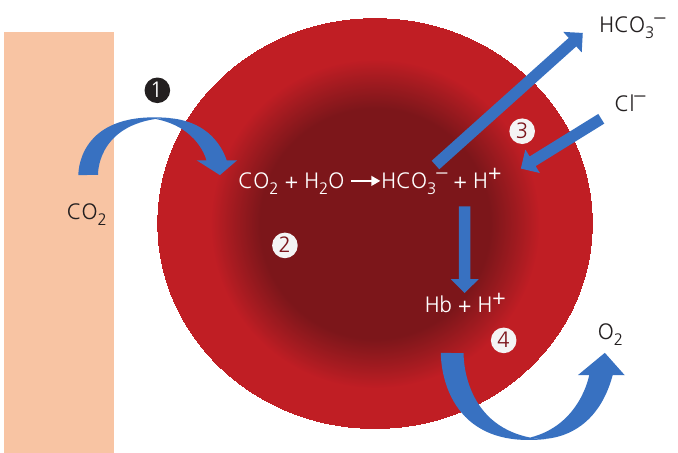
Step-by-Step: CO₂ Processing in Red Blood Cells
Step 1: Carbon dioxide enters the red blood cell
Carbon dioxide produced by respiring tissues diffuses down its concentration gradient from tissue fluid into red blood cells.
Step 2: Formation of carbonic acid
Inside the red blood cell, carbon dioxide reacts with water to form carbonic acid:
This reaction is catalyzed by the enzyme carbonic anhydrase, which is found in red blood cells but not in plasma. The enzyme dramatically increases the reaction rate.
Carbonic acid immediately dissociates into hydrogen ions and hydrogen carbonate ions:
Step 3: The chloride shift
Hydrogen carbonate ions (HCO₃⁻) diffuse out of the red blood cell down their concentration gradient into the plasma. There, they combine with sodium ions (Na⁺) to form sodium hydrogen carbonate.
To maintain electrochemical neutrality within the cell, negatively charged chloride ions (Cl⁻) diffuse from the plasma into the red blood cell, replacing the departing HCO₃⁻ ions. This exchange is called the chloride shift (also known as the Hamburger phenomenon).
Mnemonic: "CIO" = Chloride In, HCO₃⁻ Out
Step 4: Buffering by haemoglobin
The hydrogen ions (H⁺) produced when carbonic acid dissociates combine with haemoglobin (Hb) to form haemoglobinic acid:
This buffering action:
- Prevents the accumulation of H⁺ ions, which would otherwise decrease pH inside the cell
- Enhances oxygen release (contributes to the Bohr effect)
- Provides substrate for carbamino formation, allowing CO₂ to bind to haemoglobin
The binding of H⁺ to haemoglobin stabilizes the deoxyhaemoglobin form, which has lower affinity for oxygen, further promoting oxygen release in respiring tissues.
Why carbonic anhydrase is crucial:
Without carbonic anhydrase, the reaction between CO₂ and H₂O would be too slow to handle the rapid CO₂ production from cellular respiration. The enzyme speeds up this reaction by thousands of times, making efficient CO₂ transport possible.
Note that carbonic anhydrase is only present in red blood cells, not in plasma—this ensures that the formation of hydrogen carbonate ions occurs in a controlled location.
Key Points to Remember
-
Haemoglobin is a quaternary protein consisting of four polypeptide chains (2α, 2β), each with an iron-containing haem group that binds one O₂ molecule.
-
Cooperative binding creates the S-shaped oxygen dissociation curve, ensuring efficient oxygen loading in the lungs (high pO₂) and efficient unloading in tissues (low pO₂).
-
The Bohr effect occurs when increased CO₂ and decreased pH reduce haemoglobin's oxygen affinity, enhancing oxygen release in actively respiring tissues where it is most needed.
-
Fetal haemoglobin contains gamma chains instead of beta chains, giving it higher oxygen affinity to extract oxygen from maternal blood across the placenta.
-
Carbon dioxide transport occurs mainly as HCO₃⁻ ions (70–90%), with smaller amounts dissolved in plasma (5%) and bound to haemoglobin as carbaminohaemoglobin; the chloride shift maintains electrochemical balance during HCO₃⁻ formation.
-
Carbonic anhydrase in red blood cells catalyzes the rapid conversion of CO₂ to carbonic acid, which then dissociates to form HCO₃⁻ and H⁺.
-
Haemoglobin acts as a buffer by binding H⁺ ions, preventing pH changes and simultaneously promoting oxygen release through the Bohr effect.