The Roles of Enzymes (OCR A-Level Biology A): Revision Notes
The Roles of Enzymes
Introduction
Enzymes are biological molecules that make life possible. Without them, the chemical reactions necessary for life would proceed far too slowly to sustain living organisms. These remarkable proteins act as catalysts, substances that accelerate chemical reactions without being permanently altered themselves.
All enzymes share several key characteristics:
- They are globular proteins with complex three-dimensional structures
- They function in small quantities and can be reused repeatedly
- They are highly specific, catalysing only particular reactions
- They work under mild conditions (low temperatures, near-neutral pH, normal pressures)
Catalyst: A substance that increases the rate of a chemical reaction but does not become altered or changed during the reaction, so can be reused over and over again.
Enzyme: A protein molecule made by cells that acts as a catalyst and increases the rate of a chemical reaction.
Enzymes control all metabolic reactions in organisms. They build complex molecules (like starch for energy storage or collagen for structural support) and break down molecules (like during digestion). The reactions they catalyse are reversible, though enzymes do not determine which direction a reaction proceeds—only how quickly it happens.
Sites of enzyme activity
All enzymes are synthesised inside cells through the process of protein synthesis. However, their location of action varies. Some remain within cells, while others are secreted to work externally.
Intracellular enzymes
Intracellular enzymes: Enzymes that catalyse reactions within cells.
These enzymes operate exclusively inside cells, catalysing the numerous simultaneous reactions that occur in the cytoplasm and organelles. They function under relatively constant conditions: low substrate concentrations, moderate temperatures (around 37°C in mammals, ~20°C in plants), and pH close to 7.0.
A key example is catalase, which breaks down hydrogen peroxide (H₂O₂). Hydrogen peroxide forms as a waste product of metabolism and is highly toxic—it acts as a powerful oxidising agent. Catalase rapidly converts it to harmless water and oxygen:
Without catalase, hydrogen peroxide would take months to degrade naturally. With the enzyme, this occurs within seconds.
Intracellular enzymes may be:
- Free in solution (in cytosol, nucleoplasm, mitochondrial matrix, or chloroplast stroma)
- Fixed to membranes (attached to the cell surface membrane or organelle membranes)
Extracellular enzymes
Extracellular enzymes: Enzymes that catalyse reactions outside cells.
These enzymes are secreted from cells to catalyse reactions in the external environment or within body cavities. Digestive enzymes are prime examples—they break down large food molecules in the gut lumen into smaller molecules that can be absorbed.
Examples include:
- Amylase: secreted into the mouth and small intestine; hydrolyses starch into maltose
- Trypsin: secreted into the small intestine; breaks down proteins into peptides and amino acids

Some organisms rely entirely on extracellular digestion. Fungi, for instance, secrete enzymes directly onto their food source, digesting it externally before absorbing the simple molecules through their hyphal walls.
Certain digestive enzymes are fixed to cell membranes rather than free in solution. For example, sucrase is attached to the cell membranes of intestinal epithelial cells.
Mechanism of enzyme action
The active site and enzyme specificity
The remarkable selectivity of enzymes arises from the structure of their active site—a specific region of the enzyme molecule where the substrate binds. This area typically forms a groove or cleft in the enzyme's surface and comprises only a small number of amino acids from the polypeptide chain.
Specificity: The relationship between an enzyme and the only type of molecule that fits into its active site.
The active site has a shape that is complementary to the substrate molecule (not identical—this is a crucial distinction). Only substrate molecules with the correct complementary shape can fit into the active site, rather like a hand fitting into a glove.
The R groups of amino acids in the active site determine its properties:
- Non-polar R groups create hydrophobic regions that accept non-polar substrates
- Polar R groups form temporary ionic bonds with substrates
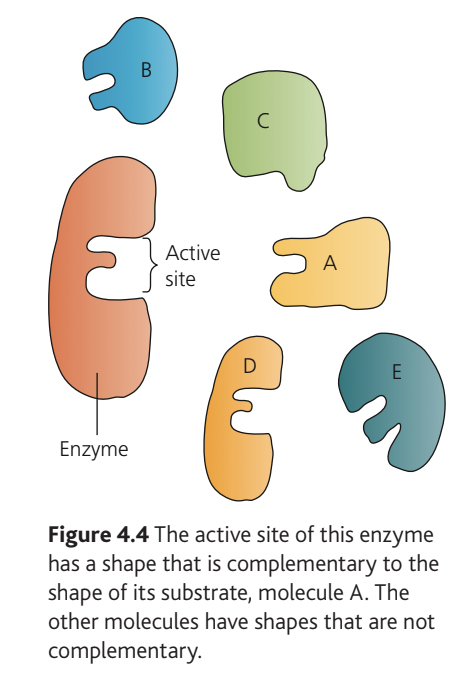
Enzymes exhibit varying degrees of specificity. Some are highly selective, catalysing only one specific reaction. Others show broader specificity, acting on a particular type of chemical bond or similar substrates.
Protease Enzyme Specificity
Consider these three protease enzymes and their specificity:
| Enzyme | Specificity (site of peptide bond cleavage) |
|---|---|
| Trypsin | Next to arginine or lysine residues |
| Chymotrypsin | Next to aromatic amino acids (phenylalanine, tyrosine, tryptophan) |
| Subtilisin | Between any pair of amino acids |
Subtilisin, used in contact lens cleaning fluid, acts as a general protease breaking peptide bonds indiscriminately. In contrast, trypsin and chymotrypsin (both secreted into the small intestine) are far more selective in their action.
Lock and key hypothesis
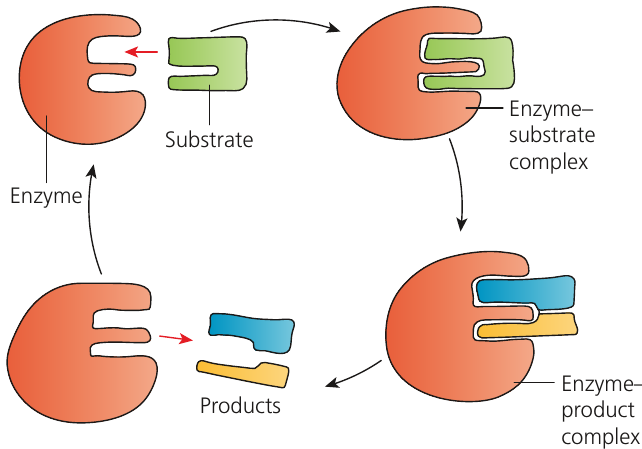
The original model of enzyme action, proposed in 1890, is the lock and key hypothesis. This suggests that the substrate fits precisely into the enzyme's active site, like a key fitting into a lock.
Lock and key hypothesis: The idea that an enzyme's active site is complementary in shape to the substrate in the same way as a key (substrate) and a lock (enzyme).
According to this model:
- The substrate enters the active site, forming an enzyme-substrate complex
- Chemical interactions between R groups in the active site and the substrate stabilise the complex
- The substrate is converted into product(s), forming an enzyme-product complex
- The product is released, leaving the enzyme unchanged and ready to accept another substrate molecule
Enzyme-substrate complex: The complex that forms when a substrate fits into an active site.
Enzyme-product complex: The complex that forms in the active site after the reaction is complete but before the product or products leave.
Induced-fit hypothesis
Research since 1890 revealed that enzyme action is more complex than the lock and key model suggests. Proteins are not rigid structures, and random collisions between enzymes and substrates would be inefficient if the active site had a fixed shape.
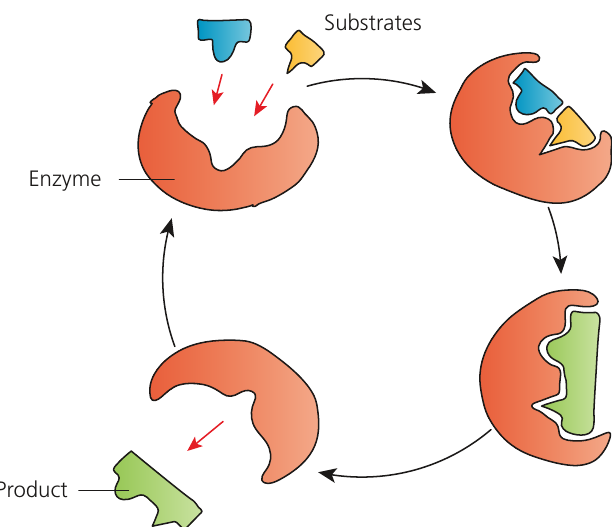
The induced-fit hypothesis, proposed in 1959, provides a more accurate model. It suggests that the active site does not have a fully complementary shape until the substrate binds. As the substrate approaches, the active site moulds around it, bringing specific R groups closer to the substrate.
Induced-fit hypothesis: The theory that the active site of an enzyme changes shape during the binding of a substrate molecule, and this puts strain on the substrate molecule contributing to the reaction.
This conformational change:
- Brings amino acids with specific R groups into optimal positions
- Holds the substrate firmly in place
- Puts strain on the substrate molecule, facilitating the reaction
- Explains how enzymes and substrates can successfully form complexes despite random collisions
Activation energy
Chemical reactions require energy to overcome an initial barrier before they can proceed. This barrier exists because the molecules involved are stable under normal conditions.
Activation energy: The energy that is needed to overcome the energy barrier to allow a reaction to occur. Enzymes provide an alternative pathway with a lower activation energy for the reaction.
Without a catalyst, hydrogen peroxide takes approximately six months to decompose, because molecules must accumulate sufficient kinetic energy to overcome the activation energy barrier. When catalase is added, the reaction is complete within seconds.
Enzymes do not alter the overall energy change of a reaction. Instead, they provide an alternative reaction pathway with a lower activation energy. This occurs because:
- The enzyme-substrate complex creates an intermediate state
- Strain within the complex (from charges on R groups and bonds between amino acids and substrate) destabilises the substrate
- The substrate more readily forms the transition state and converts to product
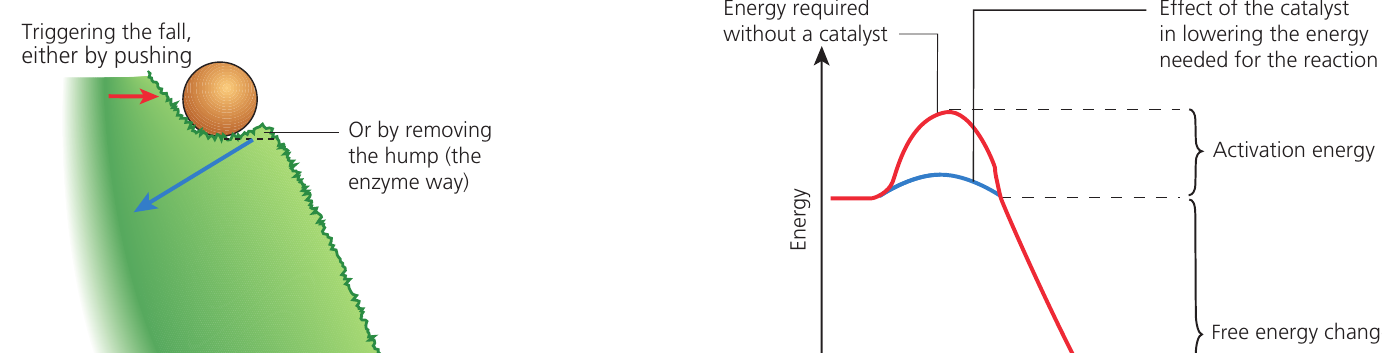
The analogy of a ball on a hillside illustrates this concept. The ball needs energy to overcome a hump before it can roll down. An enzyme effectively removes or reduces the height of that hump, allowing the reaction to proceed more easily.
Investigating enzyme activity
Enzyme activity is measured by determining the rate at which a reaction proceeds. This can be done by monitoring either:
- The disappearance of substrate
- The appearance of product
As an enzyme-catalysed reaction progresses, substrate concentration decreases. This reduces the probability of substrate molecules encountering active sites, causing the reaction rate to slow over time.
Methods for measuring enzyme activity
Following substrate disappearance: If a substrate solution is cloudy (like milk protein), you can time how long it takes for the cloudiness to disappear when a protease is added. The time taken is not the rate itself—if the reaction is fast, the time is short; if slow, the time is long. Rate is calculated as the reciprocal of time:
where = time in seconds. The unit is .
Calculating Reaction Rate
If cloudiness disappears in :
Following product appearance: When catalase breaks down hydrogen peroxide, oxygen gas is produced. This can be collected and measured using the apparatus below:
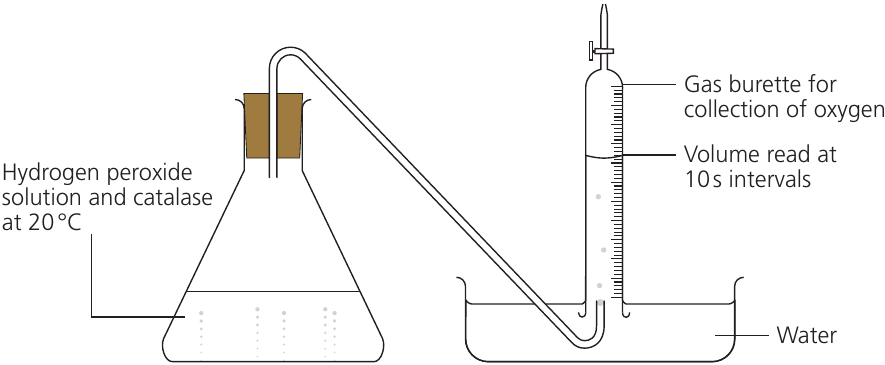
Oxygen is collected in an inverted burette or gas syringe, and the volume is recorded at regular time intervals.
Determining initial rates
When you plot the volume of product against time, you obtain a curve that starts steep and gradually levels off as substrate is depleted.
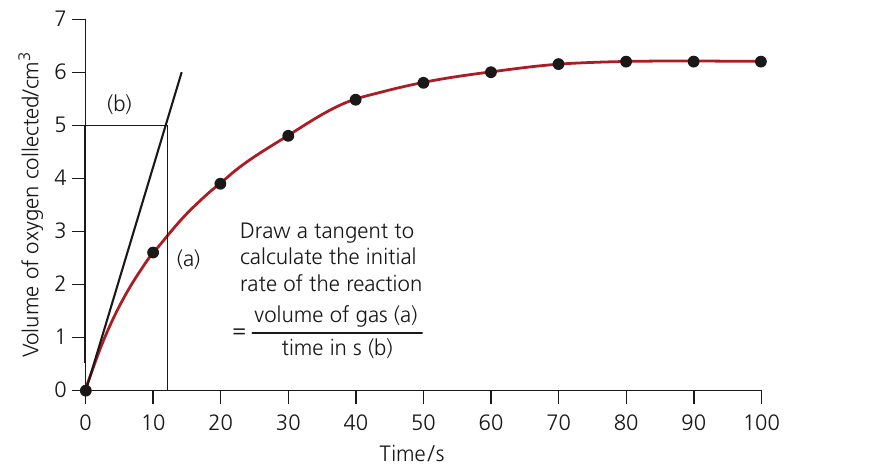
The initial rate is the rate at the start of the reaction when substrate concentration is at its maximum. This is the most reliable measure for comparing enzyme activity under different conditions because it is not affected by decreasing substrate availability.
Two methods determine initial rate:
- Calculate the rate from the first sample taken
- Draw a tangent to the curve at time zero and calculate its gradient
For the tangent method, use points separated by half the tangent's length. The gradient of the tangent gives the initial rate.
Factors affecting enzyme activity
Four main factors influence how quickly enzymes catalyse reactions: temperature, pH, substrate concentration, and enzyme concentration.
Temperature
Temperature affects the kinetic energy of molecules. Both enzyme and substrate molecules move randomly in solution, and increasing temperature makes them move faster, increasing collision frequency.
The effect of increasing temperature (below optimum):
- Molecules gain kinetic energy and move faster
- More frequent collisions occur between enzyme and substrate
- More enzyme-substrate complexes form
- Reaction rate increases
For most enzyme-catalysed reactions, the rate approximately doubles for every rise in temperature. This is expressed as the temperature coefficient, :
Temperature coefficient (): The ratio between the activities of a process at two different temperatures apart.
Calculating Temperature Coefficient
If the rate at is arbitrary units and at is arbitrary units:
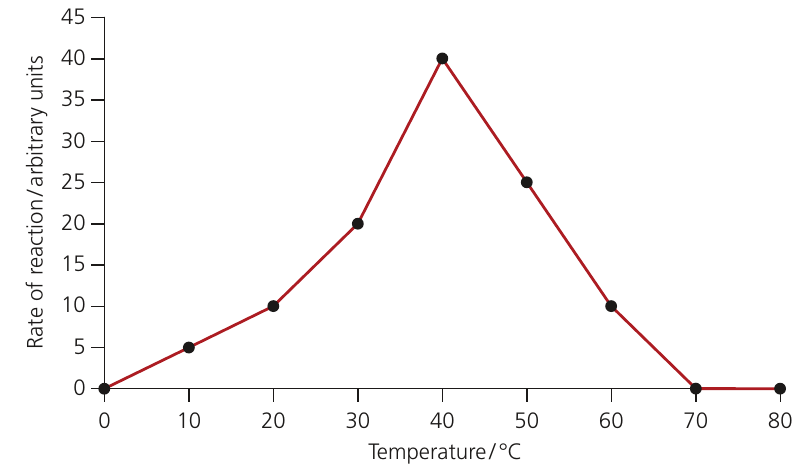
Optimum temperature: The temperature at which the rate of a process is at its maximum.
Each enzyme has an optimum temperature where it works most efficiently. This varies:
- Human and mammalian enzymes: typically (close to body temperature)
- Plant enzymes: typically around
- Thermophilic bacterial enzymes: can exceed
The effect of excessive temperature (above optimum):
At temperatures above the optimum, increased molecular vibration puts strain on the enzyme structure. The weak hydrogen bonds and ionic bonds maintaining the tertiary structure begin to break. This causes the three-dimensional shape to change, distorting the active site.
Denaturation: A change in the tertiary structure of proteins such as enzymes, which means it no longer functions.
When the active site loses its complementary shape, substrate molecules can no longer bind effectively. The enzyme is denatured—a permanent change that cannot be reversed by cooling. Note that strong covalent bonds (peptide bonds, disulfide bridges) are not broken by heat.
Most enzymes in living organisms denature above . However, enzymes from thermophilic bacteria are thermostable and can withstand temperatures of or higher. The extent of denaturation depends not only on temperature but also on exposure time—brief exposure to may leave many enzymes active, while prolonged exposure denatures most.
pH
The pH scale measures hydrogen ion () concentration. It is logarithmic, meaning each unit change represents a tenfold change in concentration. Lower pH values indicate higher concentrations (more acidic).
Each enzyme operates over a particular pH range with an optimum pH where activity is maximal. Outside this range, enzyme performance decreases, and at extreme pH values, the enzyme may not function at all.
Optimum pH is enzyme-specific:
- Gastric protease (pepsin): (highly acidic stomach environment)
- Pancreatic protease: (alkaline small intestine)
- Most intracellular enzymes: (near-neutral cytosol pH)
- Lysosomal enzymes: low optimum pH (providing safety if accidentally released into cytosol)
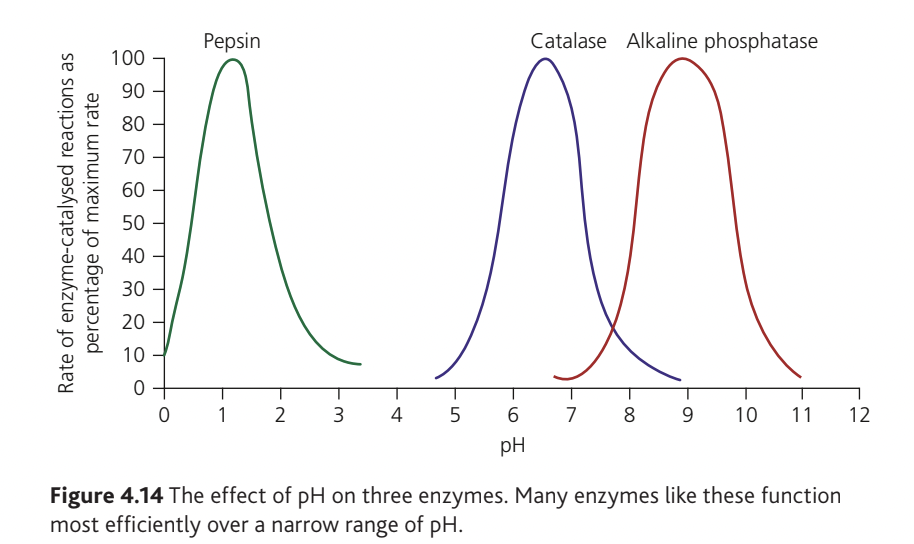
Why pH affects enzyme activity:
The active site shape depends on hydrogen bonds and ionic bonds between R groups of amino acids. Changes in concentration:
- Alter the ionisation state of R groups
- Disrupt hydrogen and ionic bonds
- Change the active site shape and charge distribution
- Reduce the ability to form enzyme-substrate complexes
Near the optimum pH, these effects are reversible. However, at pH extremes, permanent denaturation occurs. Buffer solutions are used in experiments to maintain constant pH.
Substrate concentration
Substrate concentration determines how frequently enzyme and substrate molecules collide.
At low substrate concentration:
- Few substrate molecules are present in solution
- Collision frequency between enzyme and substrate is low
- Few enzyme-substrate complexes form
- Reaction rate is slow
As substrate concentration increases:
- More substrate molecules become available
- Collision frequency increases
- More enzyme-substrate complexes form
- Reaction rate increases proportionally
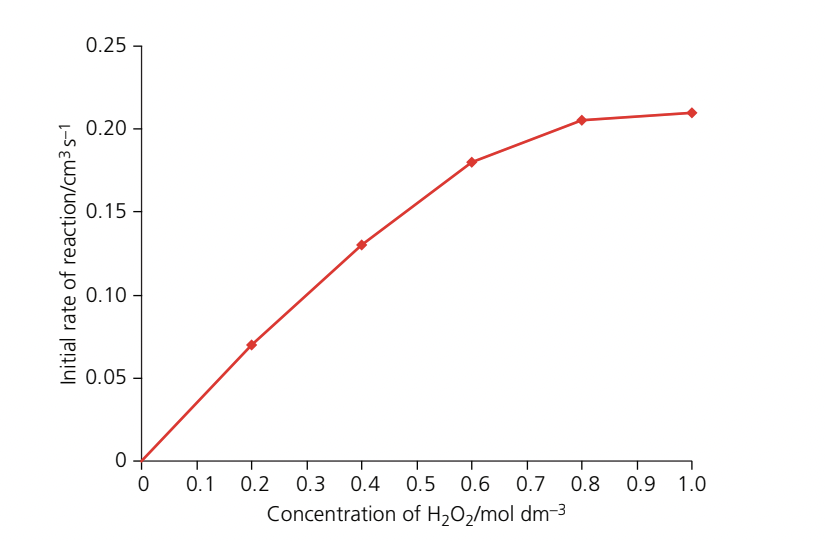
At high substrate concentration:
Eventually, a point is reached where increasing substrate concentration no longer increases the rate. The reaction reaches a maximum rate and plateaus. This occurs because:
- All enzyme active sites are occupied (saturated)
- No free enzyme molecules are available
- The rate is limited by how quickly enzymes can convert substrate to product and release it
- Further substrate increase has no effect
At saturation, the rate is limited by enzyme concentration, not substrate concentration. To increase the rate further, more enzyme would need to be added.
Enzyme concentration
When substrate is in excess, the number of enzyme molecules determines the reaction rate.
Increasing enzyme concentration:
- Provides more active sites for substrate binding
- Allows more enzyme-substrate complexes to form simultaneously
- Increases the rate of product formation
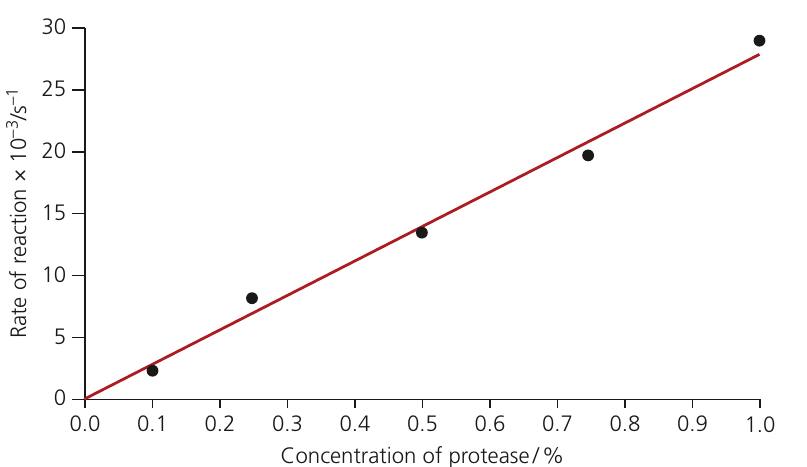
The relationship between enzyme concentration and reaction rate is linear (directly proportional) provided:
- Substrate concentration remains high (non-limiting)
- Temperature remains constant
- pH remains constant
If substrate becomes limiting, or if temperature or pH changes, these factors will influence the rate instead.
Key Points to Remember:
-
Enzymes are biological catalysts made of protein that speed up reactions without being used up, functioning in small quantities under mild conditions
-
Active sites have complementary shapes to their substrates, explaining enzyme specificity; the induced-fit model shows the active site moulds around the substrate
-
Enzymes lower activation energy by providing an alternative reaction pathway, allowing reactions to occur at body temperature
-
Four factors affect enzyme activity: temperature (rate increases to optimum then denatures), pH (each enzyme has specific optimum), substrate concentration (rate increases until saturation), and enzyme concentration (directly proportional when substrate is excess)
-
for most enzymes below optimum temperature, meaning the rate doubles for every increase