Structure and Role of Cell Membranes (OCR A-Level Biology A): Revision Notes
Structure and Role of Cell Membranes
Introduction to cell membranes
Cell membranes act as selective barriers between the cell interior and its external environment. These structures are described as partially permeable (or selectively permeable), meaning they permit certain substances to pass through whilst blocking others. This selective property allows cells to control their internal composition and maintain optimal conditions for biochemical processes.
The plasma membrane (also called the cell surface membrane) surrounds the cytoplasm and forms the outer boundary of the cell. However, eukaryotic cells also contain numerous internal membranes that enclose organelles such as mitochondria and chloroplasts, surround vacuoles, and form structures like the endoplasmic reticulum. These membranes share a similar basic structure but may vary in their protein composition depending on their specific functions.
Membranes perform three essential roles:
- Controlling substance movement through various transport mechanisms
- Cell signalling via receptor proteins that detect chemical messages from other cells
- Cell recognition through surface markers that identify cells to the immune system
The fluid mosaic model
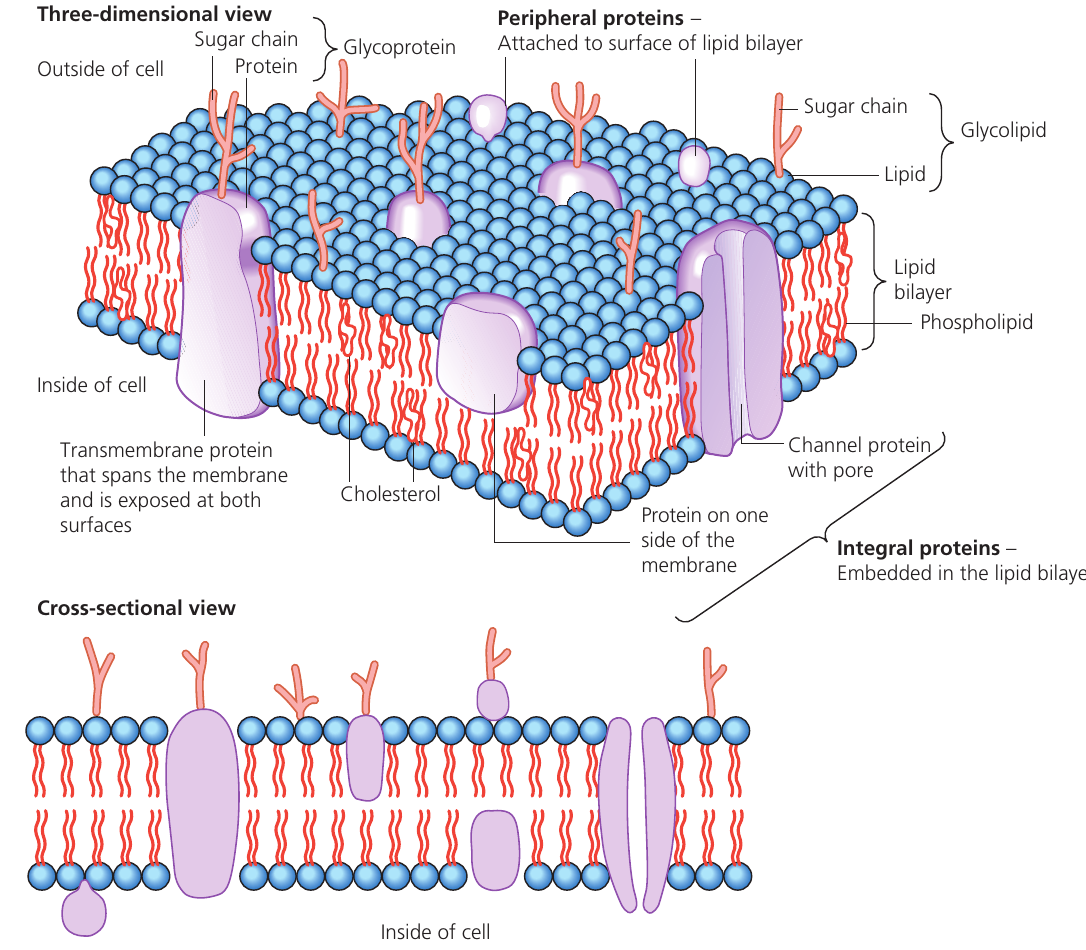
Our current understanding of membrane structure comes from the fluid mosaic model, proposed by scientists Singer and Nicolson in 1972. This model describes membranes as approximately thick structures composed primarily of lipids and proteins arranged in a specific pattern.
The term "fluid" refers to the fact that both phospholipid molecules and proteins can move laterally within the membrane layer. The term "mosaic" describes the scattered pattern of protein molecules embedded throughout the lipid structure, resembling tiles in a mosaic when viewed from the surface.
Phospholipid bilayer structure
The foundation of all cell membranes is the phospholipid bilayer, which forms the main structural component. This bilayer arrangement occurs naturally due to the unique properties of phospholipid molecules.
Phospholipid molecule structure
Each phospholipid molecule has two distinct regions:
- A hydrophilic (water-attracting) polar head containing a phosphate group
- Two hydrophobic (water-repelling) non-polar tails made of fatty acid chains
The term hydrophilic indicates an attraction to water molecules (which are also polar), whilst hydrophobic refers to a repulsion from water.
Bilayer formation
When phospholipids are placed in an aqueous environment (as exists inside and outside cells), they spontaneously arrange into a double layer. This bilayer is the most stable configuration because:
- The hydrophilic heads face outward towards the aqueous solutions (extracellular fluid outside and cytoplasm inside)
- The hydrophobic tails point inward, away from water on both sides
- The tails from each layer face each other in the membrane's centre, attracted to one another
- This arrangement minimizes unfavorable interactions between the hydrophobic tails and water
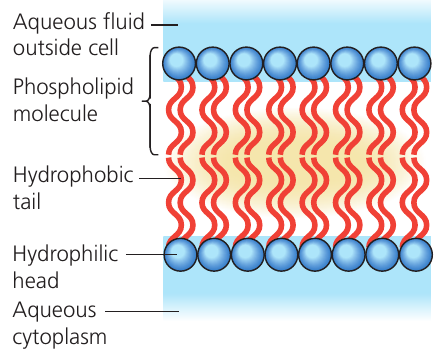
The phospholipid bilayer forms automatically in aqueous environments because the hydrophilic heads are attracted to water whilst the hydrophobic tails are repelled by it. This self-assembly process creates a stable double-layer structure where water-repelling tails are shielded from water on both sides of the membrane.
Additional stability comes from the mutual attraction between the hydrophobic fatty acid tails, which holds the bilayer together. Small amounts of glycolipids (lipids with short carbohydrate chains attached) are also scattered throughout the bilayer.
Membrane proteins
Various proteins are embedded within or attached to the phospholipid bilayer, each performing specific functions. These proteins can be classified based on their position and role.
Intrinsic and peripheral proteins
Intrinsic proteins (also called integral proteins) are embedded within the lipid bilayer. Some of these are transmembrane proteins that span the entire width of the membrane, with portions exposed on both the inner and outer surfaces.
Peripheral proteins are attached to the membrane surface rather than embedded within it. They typically associate with the heads of phospholipids or with integral proteins.
Channel proteins
Channel proteins are transmembrane proteins that form pores or passageways through the membrane. These allow water molecules and small polar (charged) substances to pass through by diffusion, moving down their concentration gradient. The substances can pass through because they travel via the protein channel rather than having to cross the hydrophobic lipid bilayer, which would normally block them.
Carrier proteins
Carrier proteins are transmembrane proteins with a different mechanism of action. These proteins can change their shape, and in doing so, they physically move substances from one side of the membrane to the other. Carrier proteins facilitate two types of transport:
- Facilitated diffusion: Moving substances down their concentration gradient (high to low concentration) without requiring energy
- Active transport: Moving substances against their concentration gradient (low to high concentration), which requires energy input
Channel proteins vs Carrier proteins:
Channel proteins form permanent pores that allow substances to pass straight through by diffusion. In contrast, carrier proteins do not form permanent pores. Instead, they bind to specific molecules, undergo a conformational change, and release the molecule on the opposite side of the membrane.
Glycoproteins
Glycoproteins are proteins with short carbohydrate (sugar) chains attached to them. These carbohydrate chains protrude from the outer surface of the plasma membrane into the extracellular environment.
Glycoproteins perform several important functions:
Membrane stabilization: The carbohydrate chains can form hydrogen bonds with surrounding water molecules, which helps stabilize the membrane structure.
Cell signalling: Glycoproteins act as receptor molecules for chemicals such as hormones. When a specific hormone binds to its receptor glycoprotein, this triggers particular changes in cellular activity. This process is called cell signalling.
Cell recognition: Glycoproteins, along with glycolipids, form surface antigens that allow the immune system to identify cells. An antigen is a molecule (typically a protein or polysaccharide) that can stimulate an immune response and trigger antibody production. The immune system uses these surface antigens to distinguish the body's own cells from foreign cells, such as those from transplanted tissue or invading pathogens.
Cholesterol in membranes
Cholesterol molecules are present in substantial quantities within the plasma membrane. Like phospholipids, cholesterol molecules have both hydrophilic and hydrophobic regions, allowing them to integrate into the membrane structure.
The primary function of cholesterol is to regulate membrane fluidity (how easily membrane components can move). Cholesterol achieves this through two mechanisms:
Preventing excessive fluidity: Cholesterol molecules bind to phospholipids, restricting their movement. This prevents the membrane from becoming too fluid, which would compromise its effectiveness as a barrier.
Preventing rigidity: Fatty acid chains in phospholipids could potentially pack together tightly and crystallize, making the membrane excessively rigid. Cholesterol molecules positioned between the fatty acid chains prevent this crystallization, ensuring the membrane remains fluid enough to function properly.
By regulating fluidity, cholesterol ensures the membrane maintains optimal flexibility for normal cellular functions whilst preserving its integrity as a barrier. It acts as a "fluidity buffer" – preventing the membrane from being either too fluid or too rigid.
Glycolipids
Glycolipids are lipid molecules with short carbohydrate chains attached. Like glycoproteins, these molecules contribute to the cell's surface antigens and play a role in cell recognition by the immune system.
Factors affecting membrane permeability
Cell membrane behavior and effectiveness as a selective barrier can be altered by environmental conditions. Two main factors affect membrane structure and permeability: temperature and organic solvents.
Temperature effects
Temperature changes affect both lipid and protein components of membranes, producing different effects at different temperature ranges.
Low to moderate temperatures: As temperature increases, the lipid component becomes increasingly fluid. This increased molecular movement reduces the membrane's effectiveness as a barrier to polar molecules, allowing some polar substances that would normally be blocked to pass through. Additionally, higher temperatures increase the rate of diffusion for any substances that can cross the membrane. These changes are reversible when temperature returns to normal.
Critical temperature (): At approximately , many membrane proteins begin to denature (lose their three-dimensional structure). Protein denaturation disrupts the membrane's organized structure, and the membrane loses its ability to function as an effective barrier. Substances can then pass freely through the damaged membrane.
Unlike the reversible fluidity changes at lower temperatures, protein denaturation is irreversible.
High temperatures: Above , membrane permeability continues to increase dramatically as more proteins denature and the membrane structure breaks down further.
Organic solvents
Most organic solvents (such as ethanol and acetone) can dissolve lipids. Since the membrane is predominantly composed of lipids, exposure to organic solvents severely disrupts membrane structure, leading to increased permeability and loss of the membrane's barrier function. This is why skin exposed to ethanol or acetone (nail varnish remover) may feel strange or damaged.
Investigating membrane permeability: the beetroot practical
The effect of temperature on membrane permeability can be demonstrated using beetroot cells. This practical investigation illustrates how environmental factors affect membrane integrity.
Beetroot cell structure
Beetroot root cells contain a large central vacuole filled with red pigmented cell sap. This pigmented solution is normally contained within the vacuole by two membranes:
- The tonoplast (the membrane surrounding the vacuole)
- The plasma membrane (the membrane surrounding the cytoplasm)
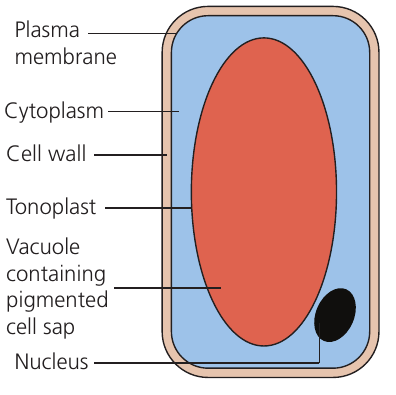
When membrane permeability increases, the red pigment leaks out from the vacuole through both membranes into the surrounding solution. The amount of pigment released can be quantified using a colorimeter, which measures how much light is absorbed by the colored solution. Higher absorbance readings indicate greater pigment release and therefore greater membrane permeability.
Experimental approach
Practical Investigation: Temperature Effects on Membrane Permeability
The investigation involves:
- Cutting uniform beetroot cylinders of equal size
- Rinsing the pieces to remove pigment from damaged surface cells
- Placing individual cylinders in tubes of distilled water at different temperatures
- Leaving them for a fixed time period
- Measuring the absorbance of the surrounding solution
Multiple repeats at each temperature allow calculation of mean values and assessment of data reliability.
Temperature effects observed
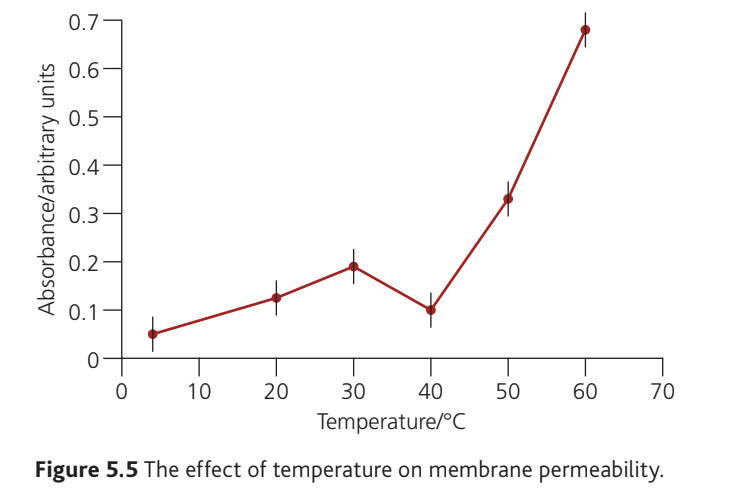
Results typically show two distinct phases:
to : Absorbance increases gradually as temperature rises. This reflects increased membrane fluidity and faster diffusion rates at higher temperatures. The membrane remains intact but becomes progressively more permeable to the pigment molecules.
to : Absorbance increases sharply and dramatically. This steep rise corresponds to protein denaturation at and above . The membrane structure breaks down, allowing much greater pigment leakage. The effect becomes more pronounced at higher temperatures as more proteins denature and membrane damage becomes more severe.
At temperatures beyond , absorbance would likely plateau once all available pigment has leaked out and/or all membrane proteins have denatured.
Remember!
Key Points to Remember:
-
The fluid mosaic model describes membranes as a phospholipid bilayer with embedded proteins that can move laterally, giving the structure both flexibility and a mosaic-like appearance.
-
Phospholipids naturally form bilayers in aqueous environments because their hydrophilic heads are attracted to water whilst their hydrophobic tails are repelled by it, creating a stable double-layer structure.
-
Membrane proteins include channel proteins (forming pores for diffusion), carrier proteins (changing shape to transport substances), and glycoproteins (functioning in cell signalling and recognition).
-
Cholesterol maintains optimal membrane fluidity by preventing both excessive fluidity and crystallization of the lipid components.
-
Temperature affects membrane permeability in two ways: gradual increases in fluidity at moderate temperatures (reversible), and protein denaturation at and above (irreversible), which severely compromises membrane integrity.