The Stages of Respiration (OCR A-Level Biology A): Revision Notes
The Stages of Respiration
Overview of the four stages
Respiration comprises four interconnected stages, each occurring in specific cellular locations:
- Glycolysis – occurs in the cytosol; does not require oxygen; functions in both aerobic and anaerobic respiration; converts glucose into pyruvate
- The link reaction – occurs in the mitochondrial matrix; converts pyruvate into an acetyl group attached to coenzyme A
- The Krebs cycle – occurs in the mitochondrial matrix; processes the acetyl group from acetyl coenzyme A
- Oxidative phosphorylation – occurs on the cristae (inner mitochondrial membranes); electrons flow along the electron transport chain, driving proton pumping and ATP synthesis
Location is Key!
Understanding where each stage occurs is crucial for understanding how respiration is organized:
- Glycolysis = Cytosol (no mitochondria needed)
- Link Reaction = Mitochondrial matrix
- Krebs Cycle = Mitochondrial matrix
- Oxidative Phosphorylation = Cristae (inner membrane)
Glycolysis
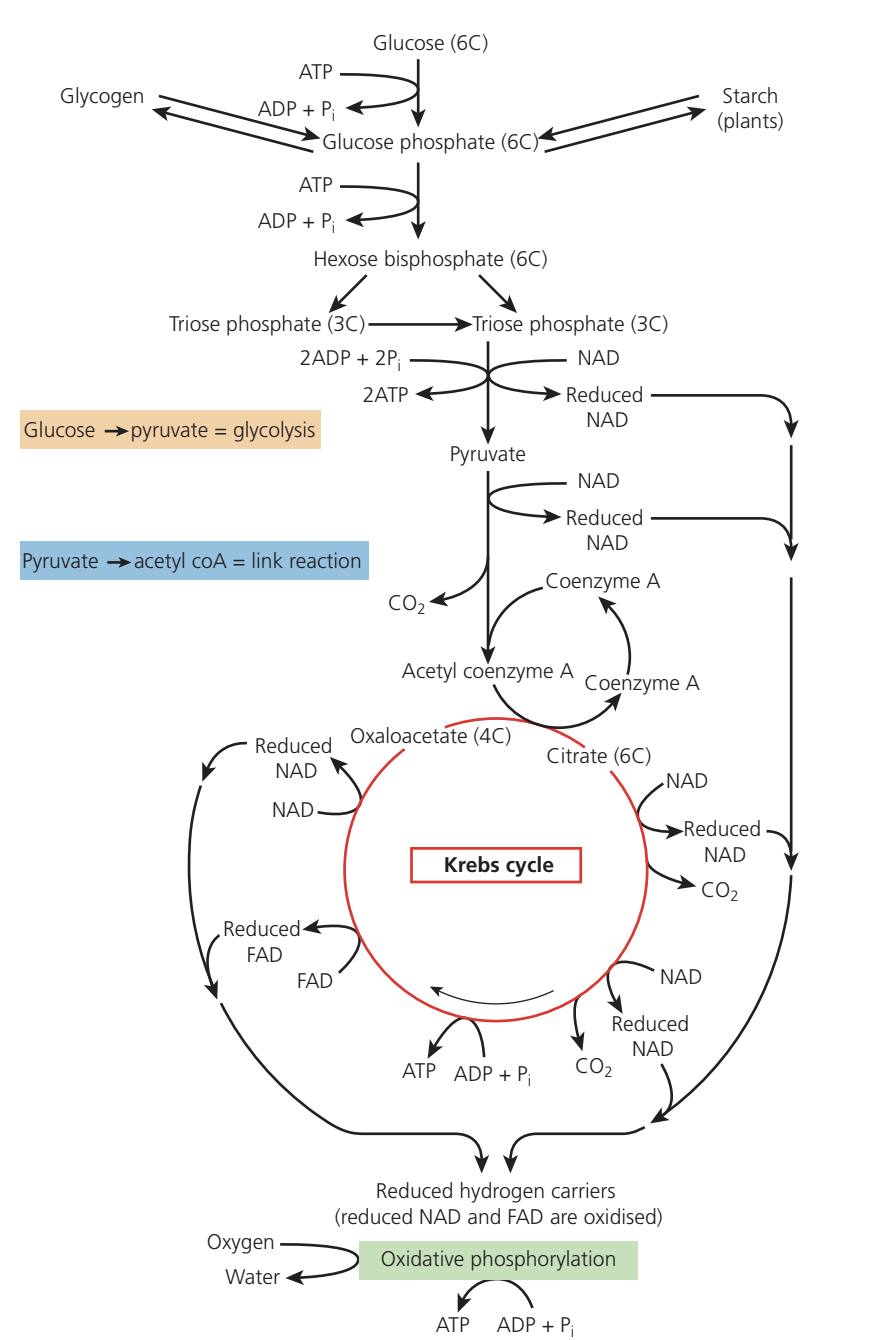
Glycolysis represents an ancient metabolic pathway that likely evolved in primitive life forms before atmospheric oxygen appeared. This anaerobic process occurs in the cytosol of all living cells and releases relatively small amounts of energy.
Ten enzyme-catalysed reactions comprise glycolysis, converting one glucose molecule into two pyruvate molecules. The pathway requires NAD coenzymes and phosphate groups from two ATP molecules.
Phosphorylation
Glucose must first be activated by phosphorylation, which raises its energy level:
- A phosphate group transfers from ATP to glucose at the carbon-6 position, forming glucose 6-phosphate
- Glucose 6-phosphate converts into its isomer, fructose 6-phosphate
- A second ATP donates another phosphate to the carbon-1 position, creating fructose 1,6-bisphosphate (the 'bis' prefix indicates phosphates attached to two different carbon atoms)
- Two ATP molecules have been consumed; the released ADP molecules will be regenerated into ATP in later stages
This initial investment of two ATP molecules is essential to "activate" glucose and make it ready for splitting. Think of it as an investment that will yield greater returns later!
Splitting fructose 1,6-bisphosphate
The six-carbon fructose 1,6-bisphosphate now splits into two three-carbon molecules, each retaining one phosphate group. These molecules are called triose phosphate (triose = three-carbon sugar).
Oxidation of triose phosphate
Dehydrogenase enzymes remove hydrogen atoms from triose phosphate molecules, with NAD acting as the hydrogen acceptor. This oxidation reaction produces:
- Two molecules of reduced NAD per glucose molecule
- Sufficient energy to add phosphate ions (from the cytosol) to both triose phosphate molecules, forming two molecules of triose bisphosphate
- Energy conservation – without this phosphorylation step, energy would dissipate as heat
Dehydrogenase is an enzyme involved in removal of hydrogen atoms from a substrate molecule. Usually associated with a coenzyme.
This is the first oxidation step in respiration where NAD is reduced!
Conversion to pyruvate
Multiple enzyme-controlled reactions convert each triose phosphate into a three-carbon pyruvate molecule. This conversion releases enough energy to phosphorylate ADP molecules directly, producing two ATP molecules through substrate-level phosphorylation – the direct formation of ATP when converting one molecule to another, contrasting with ATP formed via oxidative phosphorylation.
Products of glycolysis
Each glucose molecule entering glycolysis yields:
- Two molecules of reduced NAD – coenzymes reduced by accepting hydrogen atoms; these diffuse into mitochondria via a shunt mechanism
- Net gain of two ATP molecules – four ATP produced minus two ATP consumed in initial phosphorylation steps
- Two molecules of pyruvate – actively transported into mitochondria for the link reaction during aerobic respiration; converted to lactate (animals) or ethanol (yeast and some bacteria) during anaerobic respiration
Summary of Glycolysis:
Input: 1 glucose + 2 ATP + 2 NAD
Process:
- Phosphorylation (2 ATP invested)
- Splitting (6C → 2 × 3C)
- Oxidation (2 NAD reduced)
- Conversion to pyruvate (4 ATP produced)
Net Output: 2 pyruvate + 2 ATP + 2 reduced NAD
Location: Cytosol (no oxygen required)
The link reaction
Pyruvate molecules from glycolysis cross the mitochondrial membranes into the matrix, where enzymes for the link reaction are located. Two distinct processes occur:
- Decarboxylation – the enzyme complex pyruvate dehydrogenase removes a carboxyl group from pyruvate, releasing it as carbon dioxide ()
- Dehydrogenation – the same enzyme complex removes hydrogen atoms, which NAD accepts to form reduced NAD
The resulting two-carbon acetyl group attaches to coenzyme A, forming acetyl coenzyme A (acetyl CoA), which carries the acetyl group into the Krebs cycle.
Key Definitions:
Decarboxylation: Removal of carbon dioxide from a substrate molecule in the link reaction and in the Krebs cycle.
Substrate-level phosphorylation: Formation of ATP directly from ADP and Pi during glycolysis and the Krebs cycle when one molecule is converted to another.
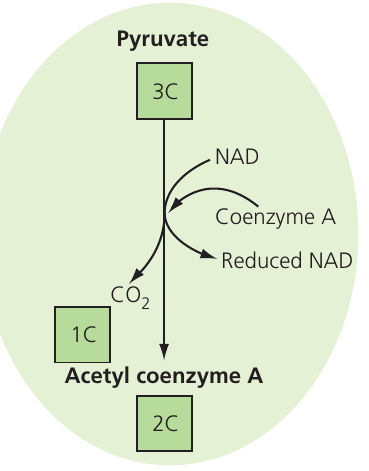
Processing two pyruvate molecules (from one glucose) through the link reaction produces:
- Two molecules of reduced NAD
- Two carbon dioxide molecules
- Two acetyl CoA molecules
The link reaction is so named because it connects glycolysis to the Krebs cycle. It's also sometimes called the oxidative decarboxylation of pyruvate because both decarboxylation and dehydrogenation occur.
The Krebs cycle
The Krebs cycle (also called the tricarboxylic acid cycle or citric acid cycle) operates in the mitochondrial matrix as a series of enzyme-controlled reactions organised cyclically. Acetyl groups from the link reaction drive this cycle.
The two-carbon acetyl group transfers from acetyl CoA to the four-carbon molecule oxaloacetate, forming the six-carbon molecule citrate. Through subsequent steps, citrate undergoes decarboxylation (releasing two molecules) and dehydrogenation (releasing eight hydrogen atoms), regenerating oxaloacetate. Six hydrogen atoms reduce NAD (forming three reduced NAD molecules), while two hydrogen atoms reduce FAD (forming one reduced FAD molecule). The energy released during these transformations directly phosphorylates one ADP to ATP through substrate-level phosphorylation.
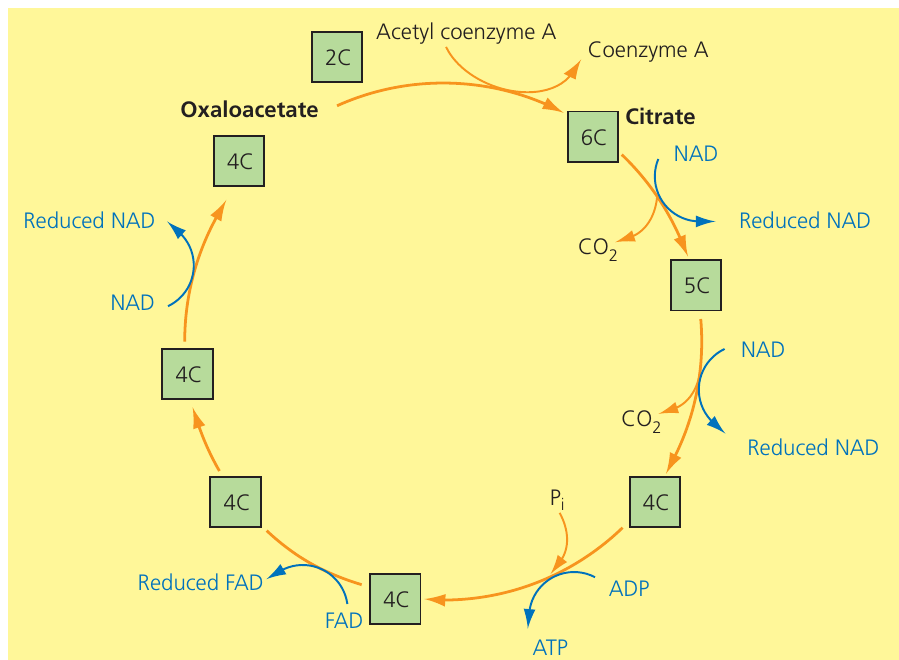
Steps in the Krebs cycle
For each turn of the cycle:
- The two-carbon acetyl group from acetyl CoA combines with four-carbon oxaloacetate, forming six-carbon citrate; coenzyme A releases for reuse
- Citrate undergoes dehydrogenation and decarboxylation, forming a five-carbon molecule; this releases hydrogen atoms (accepted by NAD, forming reduced NAD) and one molecule
- The five-carbon molecule undergoes further dehydrogenation and decarboxylation, forming a four-carbon compound plus reduced NAD and
- Reorganisation of the four-carbon molecule provides energy for direct ATP synthesis from ADP and inorganic phosphate () in the matrix – another example of substrate-level phosphorylation
- The next four-carbon compound undergoes dehydrogenation; FAD (not NAD) accepts the hydrogen atoms, becoming reduced FAD
- In the final reaction, dehydrogenation of a four-carbon molecule regenerates oxaloacetate, completing the cycle; NAD accepts the hydrogen atoms, forming reduced NAD
Products from Two Complete Turns of the Krebs Cycle (one glucose molecule):
Per turn:
- 3 reduced NAD
- 1 reduced FAD
- 2 CO₂
- 1 ATP
Total (2 turns):
- 6 molecules of reduced NAD
- 2 molecules of reduced FAD
- 4 carbon dioxide molecules
- 2 ATP molecules (produced directly)
Remember: One glucose produces two pyruvates, so two acetyl groups enter, creating two turns!
Since one glucose produces two pyruvate molecules, two acetyl groups enter the Krebs cycle, creating two complete turns. The products from two turns are:
- Six molecules of reduced NAD
- Two molecules of reduced FAD
- Four carbon dioxide molecules
- Two ATP molecules (produced directly)
Oxidative phosphorylation
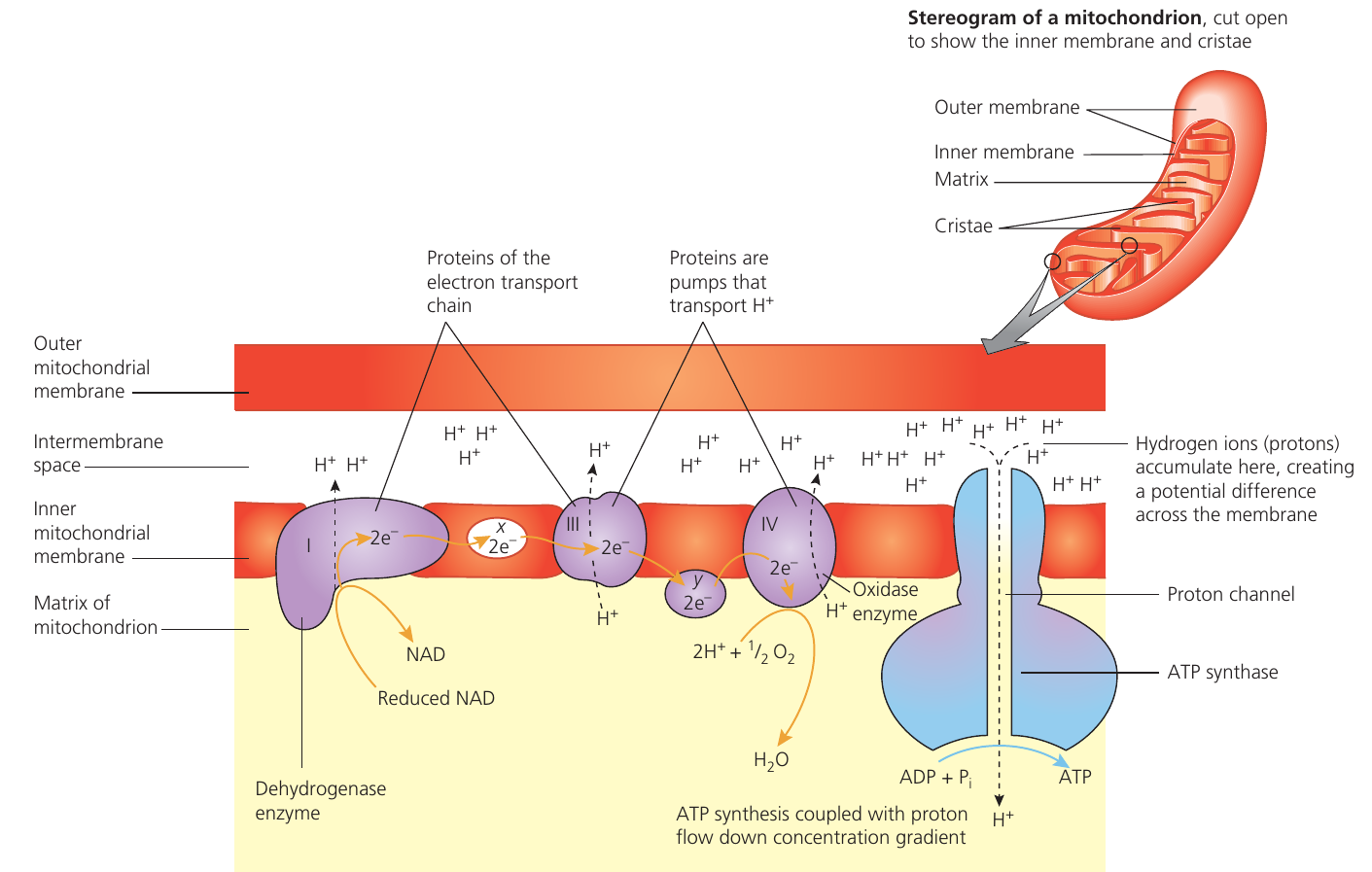
Oxidative phosphorylation uses energy from electron transfer to add inorganic phosphate to ADP, forming ATP. This process occurs on the cristae – the highly folded inner mitochondrial membranes where electron carriers embed alongside ATP synthase enzymes. The folded structure maximises surface area for carriers and ATP synthase.
Reduced NAD and FAD from the Krebs cycle (plus reduced NAD from the link reaction and possibly glycolysis) donate their hydrogen atoms to the electron transport chain. The hydrogen splits into protons () and electrons (). Electrons transfer to NADH-coenzyme Q reductase (Complex I), while protons enter the matrix.
Cristae are the highly folded inner membranes of the mitochondria where the electron carriers and enzymes are found.
The extensive folding increases surface area, allowing more electron transport chains and ATP synthase enzymes to be present, thereby increasing ATP production capacity.
Electrons pass sequentially from one carrier to another, with each carrier becoming reduced (accepting electrons) then oxidised (donating electrons to the next carrier). Energy released at certain steps phosphorylates ADP to ATP. Each pair of electrons passing through the chain produces between and ATP molecules, depending on which hydrogen carrier initiated the process and whether hydrogen atoms needed transport from the cytosol.
Oxygen serves as the final electron acceptor. It combines with electrons and protons (which rejoin the electrons), reducing to form water ().
Without oxygen as the final electron acceptor, the electron transport chain would stop functioning, as electrons would have nowhere to go. This is why oxygen is essential for aerobic respiration and why we must breathe!
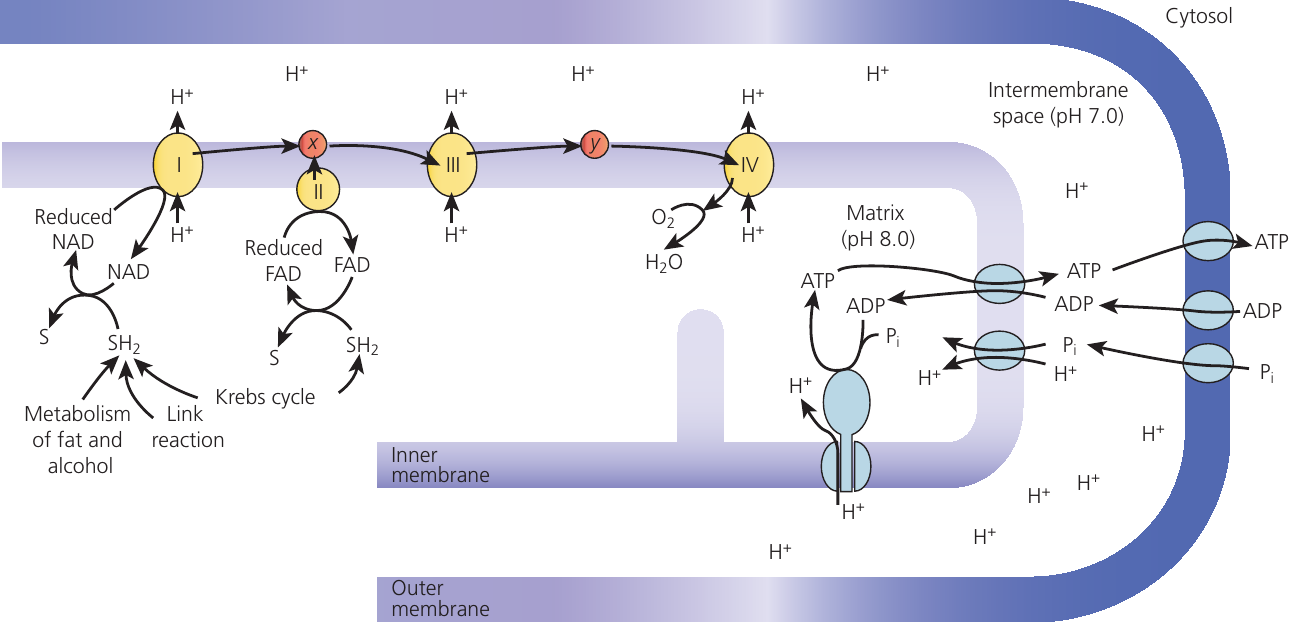
Chemiosmosis
Chemiosmosis describes ATP formation from energy released during electron transport along the electron transport chain. It occurs on the inner mitochondrial membranes as part of oxidative phosphorylation.
As electrons move along carrier molecules, they release energy. Reduced coenzymes become oxidised, and some released energy actively pumps protons into the intermembrane space. This creates a proton concentration gradient and electrochemical gradient. The pump uses energy from electron flow (electromotive force), not ATP.
Chemiosmosis: Flow of protons (hydrogen ions) through ATP synthase channels, creating an electrochemical force that is great enough to produce ATP.
This process is fundamental to ATP production in both respiration and photosynthesis!
The inner membrane remains impermeable to ions, including protons. Protons cannot cross the membrane directly but flow through ion channels associated with ATP synthase, moving down their concentration gradient. As protons flow through ATP synthase channels, the enzyme's rotating component turns, releasing energy that phosphorylates ADP with to form ATP.
The electrochemical gradient from chemiosmosis creates potential energy – stored energy from the protons' position in the intermembrane space. This potential energy drives ATP synthase and builds ATP molecules.
Chemiosmosis operates identically in photophosphorylation (photosynthesis) and occurs in mitochondrial cristae, chloroplast thylakoid membranes, and prokaryotic cell surface membranes. This similarity suggests a common evolutionary origin for these energy-conversion mechanisms.
ATP yield summary
The complete oxidation of one glucose molecule through aerobic respiration produces:
| Stage | Location | Substrate | Products | ATP yield | Respiration type |
|---|---|---|---|---|---|
| Glycolysis | Cytosol | Glucose (6C) | pyruvate (3C), ATP, reduced NAD | (net production) | Aerobic and anaerobic |
| Link reaction | Mitochondrial matrix | Pyruvate | Acetyl coenzyme A, reduced NAD, | — | Aerobic |
| Krebs cycle (per turn) | Mitochondrial matrix | acetyl groups from CoA | Oxaloacetate recycled, , ATP, reduced NAD, reduced FAD | (two turns) | Aerobic |
| Oxidative phosphorylation | Cristae | reduced NAD, reduced FAD, ADP | , ATP | – | Aerobic |
| Total | – |
Glycolysis produces four ATP molecules directly through substrate-level phosphorylation, but consumes two ATP, yielding a net gain of two ATP. The Krebs cycle produces two ATP directly (one per turn, two turns total) through substrate-level phosphorylation. Oxidative phosphorylation generates 26 ATP molecules from reduced NAD molecules.
Both reduced NAD and reduced FAD donate electrons to the electron transport chain, but only reduced NAD donates the hydrogen ions forming the proton gradient for chemiosmosis. Hydrogen ions from reduced FAD remain in the matrix, eventually recombining with electrons and oxygen to form water.
The theoretical maximum of 32 ATP is rarely achieved in practice due to:
- ATP used for active transport of pyruvate into mitochondria
- ATP used for the mitochondrial shunt mechanism transporting reduced NAD from glycolysis
- Proton leakage across the inner membrane
- Other cellular processes competing for the proton gradient
In reality, yields typically range from 30–32 ATP per glucose.
However, some ATP actively transports pyruvate into the mitochondrial matrix, and the mitochondrial shunt mechanism transports reduced NAD from glycolysis into the matrix. Some may leak out, reducing potential energy and limiting ATP production. In reality, ATP molecules may not always be achieved, with yields typically ranging from – ATP per glucose.
Key Points to Remember:
- Respiration involves four stages: glycolysis (cytosol), link reaction (matrix), Krebs cycle (matrix), and oxidative phosphorylation (cristae)
- Coenzymes NAD and FAD carry hydrogen atoms; CoA carries acetyl groups; all are essential for respiration
- Substrate-level phosphorylation (direct ATP formation) occurs in glycolysis and the Krebs cycle; oxidative phosphorylation produces most ATP
- Chemiosmosis uses the proton gradient created by the electron transport chain to drive ATP synthase
- Complete glucose oxidation yields 30–32 ATP molecules through aerobic respiration
- Oxidation = loss of hydrogen/electrons; Reduction = gain of hydrogen/electrons