Muscles and Movement (OCR A-Level Biology A): Revision Notes
Muscles and Movement
Introduction to muscle-driven movement
The brain and central nervous system coordinate all body movements through a complex network of sensory input and motor output. Movement can occur as a direct environmental response or to maintain body posture. The brain processes information from various sense organs, including stretch receptors within the muscles themselves, before sending coordinated signals to multiple muscles via motor neurones travelling through the spinal cord.
In vertebrates, movement depends on skeletal joints operated by muscles arranged in antagonistic pairs. This arrangement is necessary because muscles generate force only through contraction. Since an individual muscle cannot both flex and extend a joint, a second muscle working in opposition is required to return the joint to its original position.
Why Antagonistic Pairs?
Muscle tissue can only actively shorten (contract) - it cannot push or extend itself back to its original length. This means that while one muscle contracts to bend a joint, a second muscle working in the opposite direction is essential to straighten the joint again. This antagonistic arrangement allows for controlled, bidirectional movement at all skeletal joints.
Skeletal muscle structure
Three distinct muscle types exist in the human body: skeletal (also termed striated or voluntary), smooth (also termed involuntary), and cardiac. Skeletal muscle, responsible for moving joints, possesses a highly organized and complex structure.
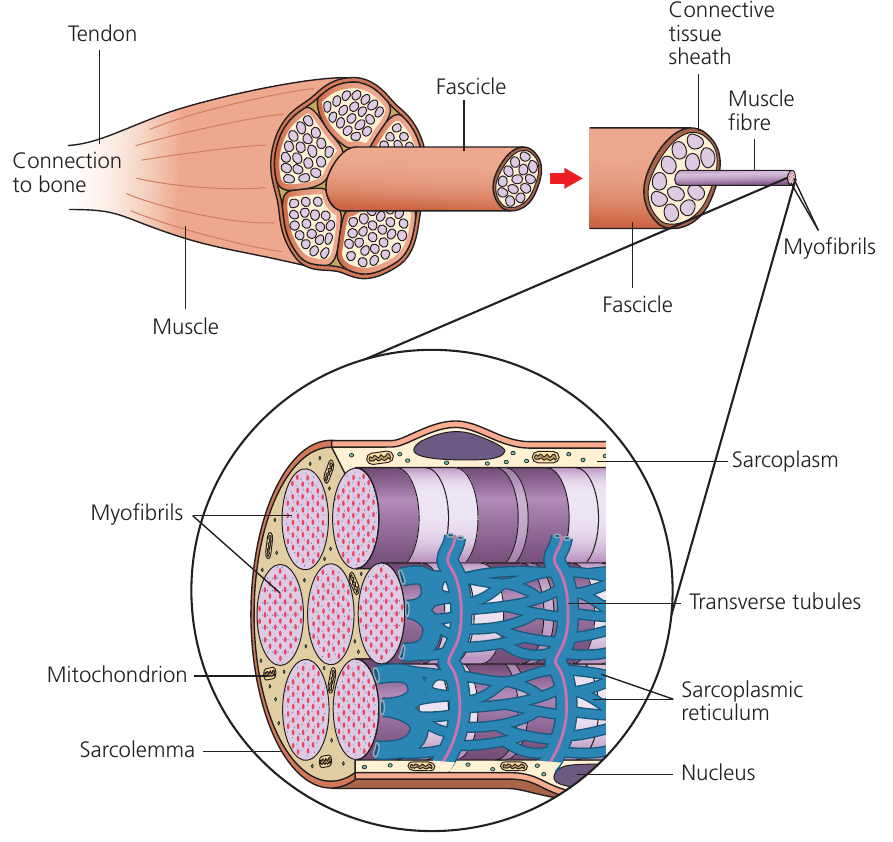
Skeletal muscle displays a hierarchical organization. Multiple muscle fibres bundle together to form a fascicle, and numerous fascicles combine to create the complete muscle. Although termed muscle "cells," these fibres are actually multinucleate structures formed through the fusion of several cells during development.

The "Sarco-" Structures
Muscle fibres contain specialized structures with the prefix sarco- (from Greek meaning "flesh"):
- Sarcolemma: the muscle fiber membrane
- Sarcoplasmic reticulum: specialized endoplasmic reticulum storing calcium ions
- Sarcoplasm: the muscle fiber cytoplasm
Think of these as specialized versions of standard cell components, adapted specifically for muscle function.
Transverse tubules (T tubules) are sarcolemma extensions that penetrate deep into the fiber, conducting electrical impulses rapidly throughout the cell. The fiber contains numerous myofibrils, which are the actual contractile elements. High ATP demand during contraction necessitates abundant mitochondria within each fiber.
Myofibril structure and the sarcomere
Myofibrils contain bundles of protein filaments that slide past one another during contraction. Two types of myofilaments exist: thick filaments composed primarily of myosin protein, and thin filaments composed primarily of actin protein. Both myosin and actin function as cytoskeletal components in many cell types.
The regular alternating arrangement of thick and thin filaments creates the characteristic striped appearance giving skeletal muscle its alternative name: striated muscle. Darker bands represent regions where filaments overlap.
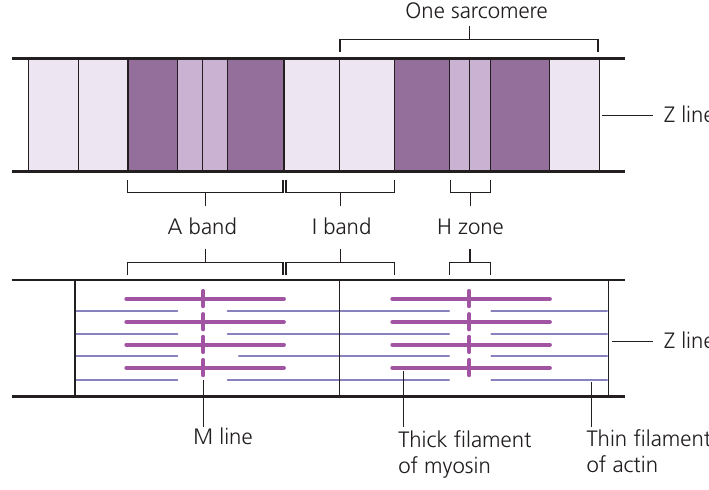
The repeating contractile unit along the myofibril is the sarcomere, bounded by Z lines at each end. Within each sarcomere:
- Z line: attachment point for thin filaments
- I band: region containing only thin filaments (appears lighter)
- A band: region containing thick filaments, including overlap with thin filaments (appears darker)
- H zone: central region containing only thick filaments
- M line: center of the sarcomere where thick filaments attach

Understanding Sarcomere Bands
The banding pattern in sarcomeres creates the "striated" appearance of skeletal muscle:
- The A band appears dark because thick and thin filaments overlap here
- The I band appears light because it contains only thin filaments
- The H zone in the center contains only thick filaments
During contraction, the I band and H zone narrow, but the A band stays the same width!
The sliding filament model
The sliding filament theory explains muscle contraction through relative movement of thick and thin filaments, which slide past each other without changing their individual lengths. Understanding this mechanism requires examination of the filament structures in detail.
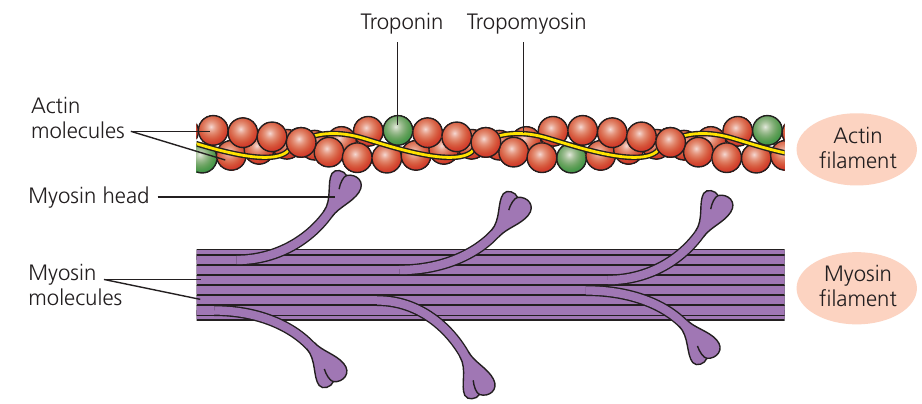
Myosin thick filaments possess projections called myosin heads capable of binding to actin and generating movement. The thin filaments contain actin molecules arranged in a twisted chain, with tropomyosin protein wound around them. Troponin protein molecules attach to tropomyosin at regular intervals.
In relaxed muscle, tropomyosin physically blocks myosin-binding sites on actin, preventing myosin heads from attaching. When calcium ions become available, they bind to troponin, causing a conformational change that moves tropomyosin away from the binding sites.
During contraction, the I band and H zone become noticeably narrower as thin filaments slide toward the sarcomere center. The A band maintains constant width because thick filament length remains unchanged.
The Role of Regulatory Proteins
Tropomyosin and troponin act as "gatekeepers" for muscle contraction:
- Tropomyosin physically covers the myosin-binding sites on actin when muscles are relaxed
- Troponin responds to calcium ions by changing shape and pulling tropomyosin aside
- This system ensures muscles only contract when calcium signals are present
Mechanism of muscle contraction
When a nerve impulse arrives at a muscle fiber, the following sequence produces contraction:
The nerve impulse depolarizes the sarcolemma. T tubules rapidly transmit this depolarization throughout the entire fiber. The depolarization triggers voltage-gated calcium channels in the sarcoplasmic reticulum to open, releasing ions into the sarcoplasm.
Calcium ions bind to troponin molecules, causing them to change shape. This conformational change pulls tropomyosin away from myosin-binding sites on actin, exposing them.
Myosin heads bind to the newly exposed sites on actin. This binding triggers release of and inorganic phosphate () already present in the myosin heads. Release of these products causes the myosin heads to bend, creating a power stroke that pulls the thin filaments toward the sarcomere center.
Fresh molecules bind to myosin heads, causing them to detach from actin. The myosin head then functions as an ATPase enzyme, hydrolyzing into and . Energy from this hydrolysis resets the myosin head to its extended position, ready to bind to the next myosin-binding site along the actin filament.
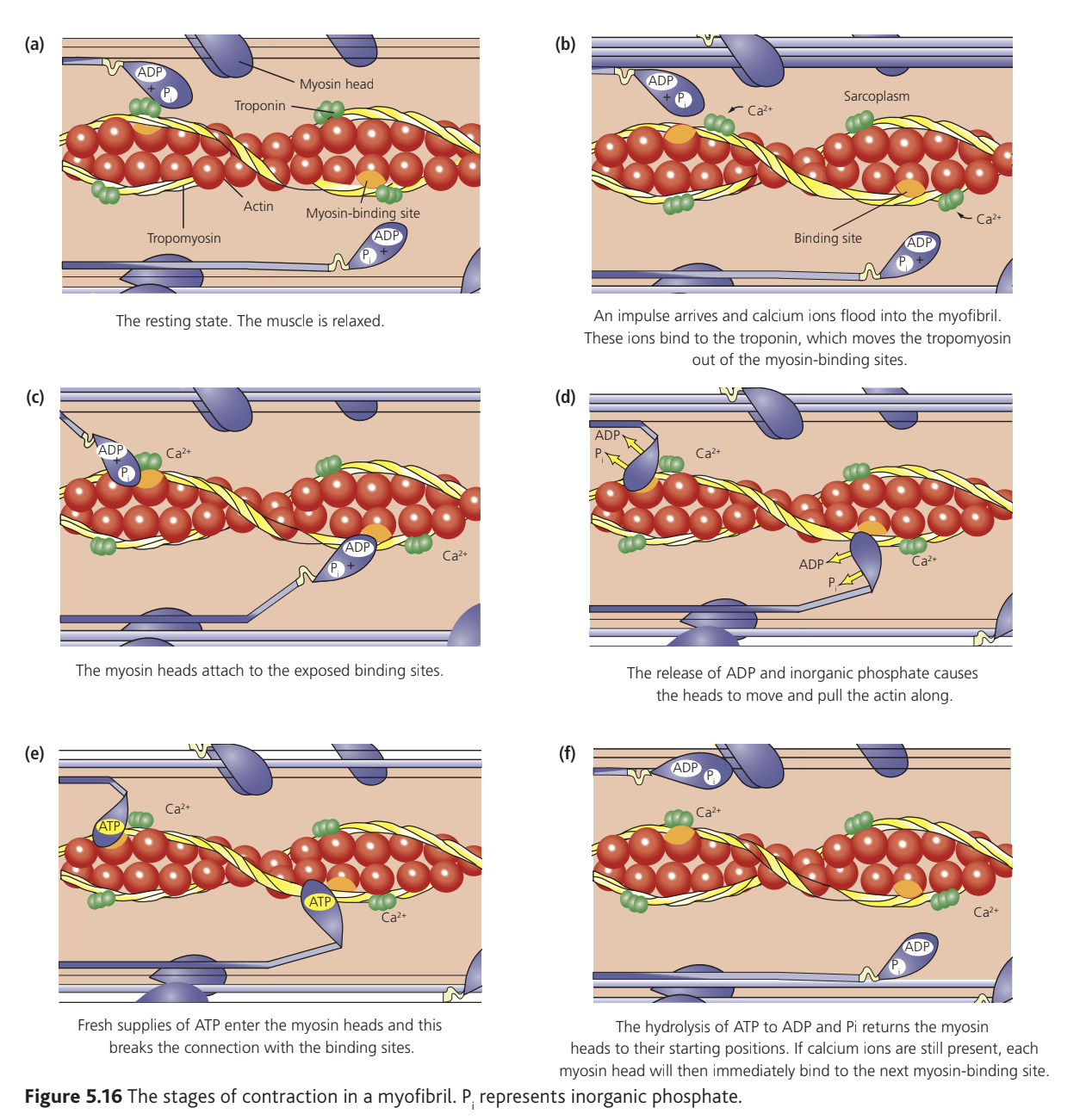
This cycle repeats continuously while calcium ions remain available, pulling thin filaments progressively toward the sarcomere center. The cumulative shortening of all sarcomeres produces contraction of the entire muscle.
When nerve impulses cease, calcium ions are actively pumped back into the sarcoplasmic reticulum. Without calcium, troponin returns to its original shape, allowing tropomyosin to block the myosin-binding sites again, and the muscle relaxes.
Worked Example: The Muscle Contraction Cycle
Let's trace one complete cycle of myosin-actin interaction:
Step 1: Calcium ions bind to troponin
- Troponin changes shape and pulls tropomyosin aside
- Myosin-binding sites on actin are exposed
Step 2: Myosin head (already charged with ADP + ) binds to actin
- Forms a cross-bridge between thick and thin filaments
Step 3: Power stroke occurs
- ADP and are released from the myosin head
- Myosin head bends, pulling actin filament toward sarcomere center
- This shortens the sarcomere
Step 4: ATP binds to myosin head
- Myosin releases from actin binding site
- Cross-bridge breaks
Step 5: ATP is hydrolyzed
- Myosin head acts as ATPase enzyme:
- Energy resets myosin head to extended "cocked" position
- Myosin head is ready to bind to the next site along actin
Result: The cycle repeats as long as calcium and ATP are available, causing progressive sarcomere shortening.
Energy supply for muscle contraction
plays an essential role in muscle contraction, with muscles requiring substantial quantities. Muscle fibers contain numerous mitochondria that normally provide sufficient through aerobic respiration. During vigorous exercise, however, supplies may become inadequate. Muscles employ several strategies to address this:
Myoglobin serves as an oxygen storage protein within muscles. This pigment releases oxygen only when surrounding oxygen concentration drops very low, providing an emergency oxygen reserve for aerobic respiration.
Myoglobin: The Muscle's Oxygen Store
Myoglobin is chemically similar to hemoglobin but has a much higher affinity for oxygen. It only releases oxygen when the partial pressure drops extremely low - essentially acting as an emergency backup system during intense muscle activity when blood supply cannot deliver oxygen fast enough.
Anaerobic respiration can support muscles for short periods during intense activity. This process provides nearly all energy for brief strenuous bursts but releases far less ATP per glucose molecule than aerobic respiration, making it inefficient for sustained activity.
The ATP-creatine phosphate system provides additional for very brief periods. Creatine phosphate () stored in muscle fibers can rapidly donate phosphate to convert into :
This reaction produces no lactate, thus avoiding muscle fatigue. However, creatine phosphate stores are extremely limited, depleting within a few seconds and providing only for very short bursts of maximum activity.
Three Energy Systems for Muscles
Muscles use three distinct energy systems depending on intensity and duration:
- Aerobic respiration - Primary system for sustained activity; requires oxygen; produces lots of ATP
- Creatine phosphate system - Immediate energy for 5-10 seconds; no oxygen needed; very limited supply
- Anaerobic respiration - Short bursts (30 seconds to 2 minutes); no oxygen needed; produces lactate and less ATP
Athletes train different systems depending on their sport: sprinters focus on creatine phosphate and anaerobic systems, while marathon runners develop aerobic capacity.
The neuromuscular junction
Skeletal muscle contraction occurs under nervous control through impulses traveling along motor neurones. Where a motor neurone meets muscle tissue, a specialized synapse called a neuromuscular junction forms.
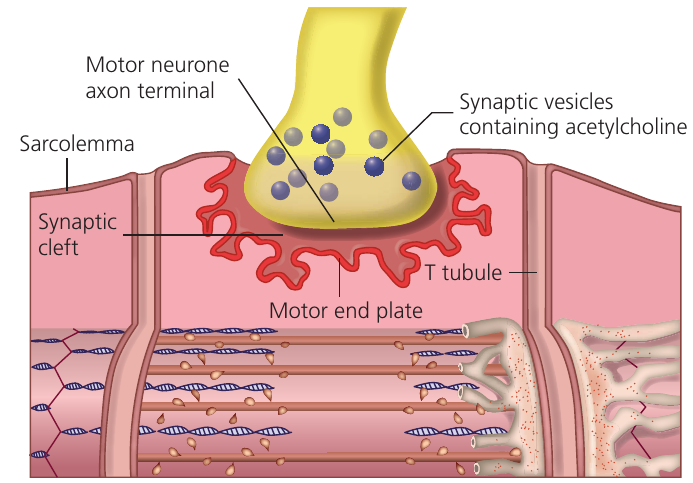
Neuromuscular junctions share similarities with nerve synapses but possess several distinctive features:
Acetylcholine functions as the neurotransmitter at all somatic motor neurones, whereas different nerve synapses employ various neurotransmitters. The postsynaptic membrane (the muscle fiber motor end plate) is extensively folded to increase surface area, allowing many more receptors than in nerve synapses. In nerve synapses, a single impulse may fail to trigger a postsynaptic action potential if threshold is not reached. In contrast, a single impulse at a neuromuscular junction invariably causes muscle fiber contraction.
Neuromuscular Junction vs. Nerve Synapse
Key differences:
- Neurotransmitter: Always acetylcholine (nerve synapses use various types)
- Postsynaptic membrane: Highly folded motor end plate with many receptors
- Response: One impulse always triggers contraction (nerve synapses may need summation)
- Reliability: Ensures consistent muscle control when needed
The neuromuscular junction operates through the following mechanism:
An impulse arriving at the motor neurone terminal causes calcium ion influx, triggering release of acetylcholine from synaptic vesicles into the synaptic cleft. Acetylcholine diffuses across the narrow cleft and binds to cholinergic receptors on the motor end plate.
This binding generates an impulse that spreads across the entire sarcolemma of the muscle fiber. Depolarization simultaneously travels down all T tubules throughout the fiber. T tubule depolarization directly couples to the contraction mechanism by opening voltage-gated calcium channels in the surrounding sarcoplasmic reticulum.
ions diffuse from the sarcoplasmic reticulum among the myofibrils, where they bind to troponin to initiate contraction. Acetylcholinesterase enzyme in the synaptic cleft then breaks down acetylcholine, preventing continuous stimulation unless additional impulses arrive.
When the sarcolemma, T tubules, and sarcoplasmic reticulum are no longer depolarized, calcium ions are actively pumped back into the sarcoplasmic reticulum, terminating contraction.
Muscle Fiber vs. Whole Muscle Contraction
An important distinction: A single impulse reaching one motor end plate always causes that specific fiber to contract. However, if only a few motor neurones send impulses, only a few muscle fibers will contract, potentially producing no noticeable movement in the whole muscle.
Muscle contraction strength depends on the number of fibers actively contracting - this is called "recruitment" and allows for fine control of movement force.
Types of muscle tissue
Fast-twitch and slow-twitch muscle fibres
Voluntary (skeletal or striated) muscles can be consciously controlled, though their actions may occur subconsciously (such as during posture maintenance). Skeletal muscles may contain one or both of two fiber types:
Fast-twitch muscle fibres contract rapidly but fatigue quickly. Slow-twitch muscle fibres contract more slowly but resist fatigue for extended periods. Some muscles consist entirely of one fiber type, while many contain both. For example, leg muscles contain slow-twitch fibers that contract continuously for posture maintenance, plus fast-twitch fibers that can be recruited for rapid leg movements.
Fiber Type Composition
Different muscles and different individuals have varying proportions of fiber types:
- Postural muscles (e.g., back muscles) contain predominantly slow-twitch fibers for sustained contraction
- Muscles for rapid movement (e.g., eye muscles) contain mostly fast-twitch fibers
- Athletic performance is partly determined by fiber type ratio - sprinters tend to have more fast-twitch fibers, while endurance athletes have more slow-twitch fibers
Smooth (involuntary) muscle
Smooth muscle (also called involuntary muscle) controls many internal organs without conscious control. It lacks the striations characteristic of skeletal muscle, though it does contain actin and myosin filaments arranged differently.
Smooth muscle forms part of the walls of various internal organs including the gut, bladder, uterus, and blood vessels. It controls processes such as gut peristalsis and arterial diameter regulation. Smooth muscle cells possess a much simpler structure than striated muscle. The cells are smaller, spindle-shaped, and contain only a single nucleus.
Cardiac muscle
Cardiac muscle exists exclusively in the heart. This specialized striated muscle possesses several unique properties:
Cardiac muscle is myogenic, meaning it contracts without requiring external nervous or hormonal stimulation. It beats rhythmically on its own, though the autonomic nervous system can regulate beating rate. Muscle fibers branch to form an interconnected network extending through atrial and ventricular walls.
Cardiac muscle never fatigues and contracts continuously throughout an organism's entire lifetime (exceeding contractions over an average human lifespan). Fibers connect to each other via specialized junctions called intercalated discs, which permit depolarization to transfer directly from cell to cell.
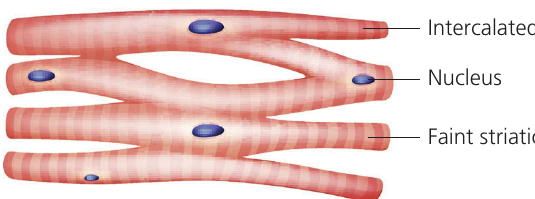
Continuous cardiac muscle contraction requires substantial energy, so cells are densely packed with mitochondria to generate the necessary .

Why Cardiac Muscle Never Fatigues
Several features allow cardiac muscle to contract continuously for a lifetime:
- Extremely high mitochondrial density - up to 40% of cell volume
- Myogenic contraction - no dependence on external signals that might fail
- Intercalated discs - ensure synchronized contraction across entire heart chambers
- Rich blood supply - constant oxygen and nutrient delivery
- Efficient aerobic metabolism - minimal lactate production
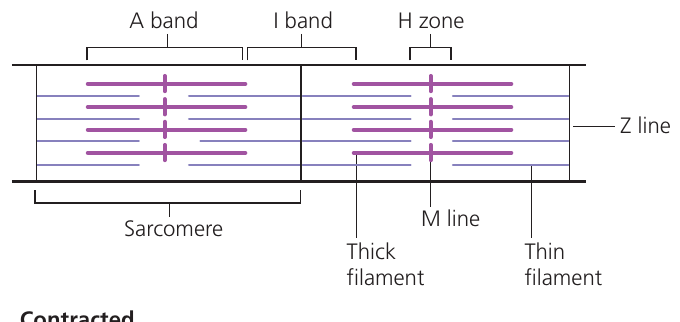
Summary diagram
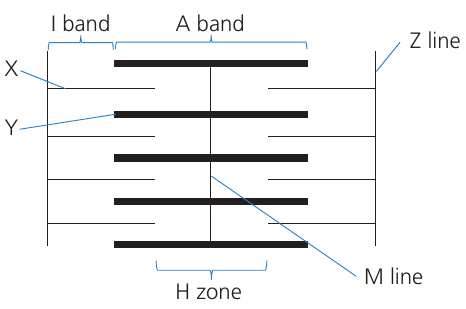
This sarcomere diagram illustrates key structures in relaxed muscle: Z lines define sarcomere boundaries, the A band contains thick filaments, I bands contain only thin filaments, and the H zone contains only thick filaments in the sarcomere center.
During contraction, the H zone and I band narrow as thin filaments slide toward the M line. The A band width remains constant because thick filament length does not change.
Key Points to Remember:
-
Muscles work in antagonistic pairs because they can only generate force through contraction, requiring opposing muscles for movement in both directions.
-
The sarcomere is the functional contractile unit, containing thick myosin and thin filaments that slide past each other without changing length.
-
Calcium ions trigger contraction by binding to troponin, which moves tropomyosin to expose myosin-binding sites on actin.
-
ATP provides energy for both the power stroke and myosin head detachment; myosin heads function as ATPase enzymes to reset for the next cycle.
-
Three energy systems support muscle activity: aerobic respiration (primary), the creatine phosphate system (rapid, brief), and anaerobic respiration (short bursts with lactate production).
-
A neuromuscular junction always produces contraction in response to a single impulse, unlike nerve synapses which may require summation to reach threshold.
-
Cardiac muscle is unique in being myogenic (self-contracting), never fatiguing, and connected by intercalated discs that allow electrical coupling between cells.